

Introduction
Numerous studies have emphasized the importance of physical activity in maintaining and improving skeletal integrity, with a well-documented relationship between exercise and bone health. The World Health Organization (WHO) advises individuals to participate in a minimum of 150 minutes of moderate-intensity aerobic exercise or 75 minutes of vigorous-intensity aerobic exercise each week to enhance overall health. The World Health Organization recommends engaging in weight-bearing endurance and resistance activities for 30-60 minutes per session, three to five days a week, to specifically enhance bone mass density.1
Physical activity affects bone health through multiple pathways, such as the mechanical stresses produced by muscle contractions, the gravitational pressure on bones, and the impact of different endocrine and paracrine variables. Exercise-induced mechanical stress induces bone remodelling by enhancing the function of osteoblasts and osteoclasts, resulting in heightened bone strength and density. Maintaining bone health and preventing disorders like osteoporosis, characterized by decreased bone mass and increased susceptibility to fractures, depends on this process.2
In addition to physical activity, the importance of dietary supplements, such as antioxidants like vitamins C and E, has gained widespread recognition. These vitamins are vital for a range of biochemical and physiological processes, but the human body is unable to produce them internally; hence, it is necessary to obtain them through diet.3 Vitamin C, a powerful antioxidant, functions as an electron donor and a reducing agent. It has a significant role in the production of collagen, which is essential for bone matrix formation. Osteogenic growth factors, such as transforming growth factor (TGF)-β, estrogen receptor (ER)-α, and osteopontin (OPN), are controlled by vitamin C. These factors then impact the formation of osteoclasts and the breaking down of bone. Research in epidemiology has demonstrated a correlation between a lack of vitamin C and a higher likelihood of developing osteoporosis and experiencing fractures. This is because a lack of vitamin C impedes bone building.4
Similarly, vitamin E, a potent antioxidant, hinders the generation of reactive oxygen species (ROS) during lipid oxidation and impedes the progression of free radical reactions. Vitamin E improves bone health by promoting the growth of trabecular bone, protecting against oxidative stress-induced bone calcium depletion, and preventing bone calcium loss in experimental models of bone loss, like ovariectomized rats.5 Antioxidant capabilities of vitamins C and E are particularly beneficial in mitigating oxidative stress resulting from intense physical activity. This stress can cause cellular harm, including bone cell damage.6
The demonstrated benefits of vitamins C and E in lowering oxidative stress and facilitating muscle recovery after exercise are well-known. However, their precise impact on bone cellular adaptability, especially in the setting of high-intensity exercise, is still uncertain. Previous studies primarily examined the impact of these vitamins on muscular performance and recuperation, leading to a limited understanding of their effects on bone health.7,8 A previous in vitro study revealed that vitamin C positively influences osteoblast proliferation in bovine. However, Vitamin E does not seem to affect osteoblast proliferation.9 Another study also showed no difference in the bone healing rate of rats treated with a combination of Vitamin C and E.10
The objective of this study is to examine the impact of high-intensity exercise, together with the addition of vitamin C and E supplements, on the process of bone cellular adaptation in mice. The research aims to investigate the potential synergistic advantages of combining antioxidant supplementation with exercise to improve bone health.
Materials and Methods
Criteria and sample group distribution
The study involved doing experimental research on a sample of 28 female mice (Mus musculus) obtained from the Farma Veterinary Center (PUSVETMA) in Surabaya. The mice selected had to be healthy and not pregnant. The mice were between the ages of 3 – 4 months, and weigh between 18 – 35 grams. They were then categorized into four distinct groups:
Control group (CON) receiving neither vitamin C nor exercise intervention.
Group 1 (VIT ) was administered a mixture of vitamins E and C only.
Group 2 (EXC) exclusively received the swimming exercise treatment.
Group 3 (CBD) received combined exercise and vitamin treatment.
The research was conducted at the Embryology Laboratory of the Faculty of Veterinary Medicine, Airlangga University, Surabaya. The mice were kept in similar cages, with diet comprising of pellet feed and distilled water.
Determination of doses of vitamin C, E, and exercise
The dosages of Vitamin C and E were adjusted from human consumption levels to suitable levels for mice.
Table 1: Dosage conversion from Human to Mice
| Mice 20 gram |
Mouse 200 gr |
Guinea Pig 400 gr |
Rabbit
1.5 kg |
Cat 2 kg |
Monkey 4 kg |
Dog 12 kg |
Human 70 kg |
|
|
Mice |
1 | 7 | 12.25 | 27.8 | 29.7 | 64.1 | 124.2 | 387.9 |
| Mouse 200g |
0.14 | 1 | 1.74 | 3.9 | 4.2 | 9.2 | 17.8 |
56 |
|
Guinea Pig |
0.08 | 0.57 | 01.00 | 2.25 | 2.4 | 5.2 | 10.2 | 31.5 |
| Rabbit 1.5kg | 0.04 | 0.25 | 0.44 | 1 | 1.08 | 2.4 | 4.5 |
14.2 |
| Cat 2kg | 0.03 | 0.23 | 0.41 | 0.92 | 1 | 2.2 | 4.1 |
13 |
|
Monkey |
0.016 | 0.11 | 0.19 | 0.42 | 0.45 | 1 | 1.9 | 6.1 |
|
Dog |
0.008 | 0.06 | 0.1 | 0.2 | 0.24 | 0.52 | 1 |
3.1 |
| Human 70 kg |
0.0026 | 0.018 | 0.031 | 0.07 | 0.076 | 0.16 | 0.32 |
1 |
From the conversion table (Laurence & Bacharach, 1964), the conversion value from humans to mice is 0.0026 / 20 g BW. The result of converting a dose of 1000 mg of vitamin C in humans is 0.13 mg/gr BW, while the result of converting 400 IU of vitamin E from the human dose is calculated by first adjusting the value of 1 International Unit (IU) to 0.67 mg so that the final value of the dose for mice is 0.03484 mg/gr BW.11 High-intensity training, using 18% body weight, involves training for 90% of the maximum time (T90) in two intervals, with a rest period between each interval equal to twice T90. Training at a frequency of 3 times per week for 28 days was shown to reduce levels of bone formation (osteoprotegerin) and increase levels of bone resorption (c-telopeptide).12 The load was attached at a point 1 cm from the base of the tail, with two intervals and rest periods of 2 times the time it takes for 90% of the load to be completed (T90), carried out three times a week for a duration of 28 days. The maximum duration was measured from the time the mice were introduced into the pool until they became unable to swim, as evidenced by sinking and the emission of substantial air bubbles.
Histology Analysis
Following the treatment period, femur bones were gathered from the mice to conduct an examination of the number of osteoblasts, osteocytes, and osteoclasts. A third-party laboratory performed the sample processing and analysis. Paraffin blocks were used to make histological preparations of femur bones. These blocks were first decalcified using a solution containing 7 grams of aluminum chloride (Al Cl3), 5 ml of formic acid, 8.5 ml of HCl, and 100 ml of distilled water. The solution was rendered inert by adding 2% sodium sulfate (Na2SO4) and left for 24 hours. The solution was then replenished multiple times to eliminate the impact of decalcification. Subsequently, the samples were rinsed with water for a duration of 12 hours. The femur metaphysis was stained using hematoxylin-eosin (HE) on longitudinal cross-sectional slices.13,14
The research findings were examined by the research team, comprised of multiple specialists in the field of histology. The histological preparations were captured using a DS Fi2 300-megapixel digital camera attached to a Nikon H600L light microscope. Histological observations of osteoblast, osteocyte, and osteoclast expressions were conducted at a magnification of 400x using the Image-J System image processing program.
Data Analysis
The descriptive data were presented as the mean (standard deviation) and were visually represented using boxplots. The Shapiro-Wilk and Levene’s test was used to determine the distribution of the data. If the data follows a normal distribution, an analysis of variance (ANOVA) was performed to assess the variations among the four groups. If the data was found to follows a normal distribution, an analysis of variance (ANOVA) was performed to assess the variations among the four groups. If the data did not follow a normal distribution, the Kruskal-Wallis test was employed as an alternative to ANOVA. Fisher’s Least Significant Difference (LSD) test was employed for post-hoc examination of regularly distributed data, whereas the Mann-Whitney test was utilized for non-normally distributed data. The statistical analyses were conducted using R studio with R version 4.3.2.
Results
The effect of Vitamin C and E supplementation in conjunction with high-intensity exercise on bone cellular adaptability are detailed in Table 2 and illustrated in Figure 1, 2, and 3.
Table 2: The average number of osteocyte cells, osteoblasts, osteoclasts, and thickness of the femoral bone cortex in each group.
|
Group |
Osteocyte
(Σ cell) |
Osteoblast
(Σ cell) |
Osteoclast
(Σ cell) |
Cortex Thickness(μm) |
|
CON |
50.14 ± 4.842 | 33.14 ± 2.755 | 1.57 ± 0.202 | 117.54 ± 4.9981 |
| VIT | 55.14 ± 3.019 | 37.43 ± 3.123 | 1.00 ± 0.000 |
118.358 ± 3.4595 |
|
EXC |
26.86 ± 4.667 | 25.86 ± 4.667 | 1.29 ± 0.184 | 121.098 ± 12.3542 |
| CBD | 54.7 ± 7.154 | 37.86 ± 3.635 | 0.714 ± 0.184 |
118.524 ± 4.5371 |
| (a) Data is displayed in mean±SD; n=7 for each group. | ||||
In Table 2, a considerable variation in the average number of osteocyte cells among the groups. The VIT group exhibited the largest mean number of osteocytes (55.14 ± 3.019 cells), with the CBD group closely following (54.7 ± 7.154 cells). The control group had a mean of 50.14 ± 4.842 cells, whereas the EXC group had the lowest count of 26.86 ± 4.667 cells.
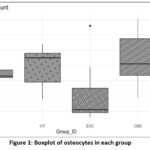 |
Figure 1: Boxplot of osteocytes in each group |
Figure 1 displays the distribution of osteocyte numbers among the various groups. The boxplot indicates that the CBD group has a larger range of osteocyte counts, ranging from 45 to 65 cells, suggesting a high degree of variability within this group. The EXC group demonstrates the lowest cell counts, predominantly falling within the range of 20 to 35 cells. The control and VIT groups have more concentrated distributions, with median values approximately at 50 and 55 cells, respectively. The histograms reveal that the control group exhibits a peak at approximately 50 cells, the VIT group shows a peak at roughly 55 cells, the EXC group displays a peak at approximately 30 cells, and the CBD group demonstrates a rather even distribution across the whole range.
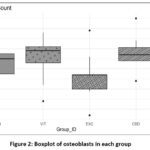 |
Figure 2: Boxplot of osteoblasts in each group |
There was variation in the quantity of osteoblasts among the different groups. The CBD and VIT groups exhibited comparable average osteoblast counts (37.86 ± 3.635 cells and 37.43 ± 3.123 cells, respectively), which were both higher than the control group (33.14 ± 2.755 cells) and considerably higher than the EXC group (25.86 ± 4.667 cells).
Figure 2 displays the distribution of osteoblast numbers. The boxplots demonstrate that the CBD and VIT groups have narrow interquartile ranges, suggesting a higher level of consistency in osteoblast counts around the median value of 38 cells. The EXC group exhibits the lowest cell counts, ranging from 20 to 30 cells, whereas the control group had a median of approximately 33 cells. The histograms demonstrate that the control group has a peak at approximately 33 cells, both the VIT and CBD groups display peaks around 37 cells, and the EXC group exhibits a wider, lower distribution with a peak around 25 cells.
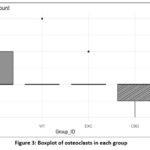 |
Figure 3: Boxplot of osteoclasts in each group |
The control group had the highest mean number of osteoclasts (1.57 ± 0.202 cells), while the EXC group had a slightly lower mean number of osteoclasts (1.29 ± 0.184 cells). The VIT and CBD groups had the lowest osteoclast counts, with mean values of 1.00 ± 0.000 cells and 0.714 ± 0.184 cells, respectively.
Figure 3 illustrates the dispersion of osteoclast numbers. The control group exhibits the greatest frequencies within a limited range, indicating a high level of stability around the mean value of 1.57 cells. The EXC group has a little wider spectrum, although still demonstrates lower frequencies in comparison to the control group. Among all the groups, the CBD group has the most decreased osteoclast counts, while the VIT and CBD groups have the lowest counts.
 |
Figure 4: Boxplot of femoral cortex thickness in each group |
The femoral cortex had a constant thickness across all groups, with no statistically significant differences seen (p≥0.05), as shown in Table 2. The group that solely performed exercise (EXC) had the highest average thickness (121.098 ± 12.3542 μm), whereas the control group had the lowest average thickness (117.54 ± 4.9981 μm). The groups that received vitamin supplementation (VIT and CBD) had comparable cortical thicknesses, measuring 118.358 ± 3.4595 μm and 118.524 ± 4.5371 μm, respectively.
Table 3: ANOVA test of the three groups (CON, EXC, VIT, CBD)
| F Value | p-Value | Effect Size | Lower CI | |
|
Osteocyte |
3.49 | 0.029* | 0.27 |
0.02 |
| Osteoblast | 1.90 | 0.152 | 0.17 |
0.00 |
(a) * significantly different (p<0.05)
Table 4: Fischer’s Least Significant Difference Post Hoc Test for osteocyte cells
|
T-Ratio |
p-Value | Estimate | SE | df |
Cohen d |
|
|
CON-EXC |
1.45 | 0.157 | 11.44 (-4.69, 27.58) | 7.88 | 28 | 0.76 (-0.23, 3.87) |
| CON-VIT | -0.93 | 0.360 | -7.33 (-23.47, 8.8) | 7.88 | 28 |
-0.73 (-2.51, 0.24) |
|
CON-CBD |
-1.40 | 0.172 | 11.71 (-5.4, 28.83) | 8.36 | 28 | 0.81 (-0.25, 2.29) |
| EXC-VIT | -2.55 | 0.017* | -18.78 (-33.87, -3.68) | 7.37 | 28 |
-1.14 (-3.82, -0.1) |
|
EXC-CBD |
-2.94 | 0.006* | 23.16 (7.02, 39.3) | 7.88 | 28 | 1.19 (0.18, 3.51) |
| VIT-CBD | -0.56 | 0.582 | 4.38 (-11.76, 20.52) | 7.88 | 28 |
0.27 (-1.07, 1.51) |
(a) * significantly different (p<0.05)
The osteocyte counts were found to have a normal distribution (p = 0.50) and homogenous variance (p = 0.43). The ANOVA analysis revealed a statistically significant effect of the group on osteocyte counts (F(3,28) = 3.492, p = 0.0286). Table 4 shows the post hoc LSD indicated that the exercise-only group (EXC) had a substantially lower number of osteocytes compared to the control group (p = 0.017), the group receiving only vitamin supplements (VIT) (p = 0.006), and the combination group (CBD). The EXC group exhibited a significant decrease in osteocyte counts compared to both the VIT group (Estimate = -18.78, SE = 7.37, p = 0.017) and the CBD group (Estimate = -23.16, SE = 7.88, p = 0.006). There were no statistically significant differences observed between the control group and the other groups
Table 5: Fischer’s Least Significant Difference Post Hoc for osteoblast cells
|
|
T-Ratio | p-Value | Estimate | SE | df | Cohen d |
| CON-EXC | 1.09 | 0.287 | 5.67 (-5.019, 16.35) | 5.22 | 28 |
0.55 (-0.43, 2.17) |
|
CON-VIT |
-0.62 | 0.541 | -3.22 (-13.908, 7.46) | 5.22 | 28 | -0.33 (-1.65, 0.74) |
| CON-CBD | -1.06 | 0.299 | 5.86 (-5.477, 17.19) | 5.53 | 28 |
0.64 (-0.54, 1.76) |
|
EXC-VIT |
-1.82 | 0.079 | -8.89 (-18.885, 1.11) | 4.88 | 28 | -0.79 (-2.83, 0.13) |
| EXC-CBD | -2.21 | 0.035* | 11.52 (0.838, 22.21) | 5.22 | 28 |
1.08 (0.2, 2.66) |
|
VIT-CBD |
-0.50 | 0.617 | 2.63 (-8.051, 13.32) | 5.22 | 28 |
0.26 (-0.96, 1.23) |
(a) * significantly different (p<0.05)
Osteoblast counts were found to follow a normal distribution (p = 0.70), and had a homogeneous variance (p = 0.97). The ANOVA analysis revealed that there were no significant differences across groups for osteoblast counts (F(3,28) = 1.902, p = 0.152). Table 5 show the pairwise post hoc LSD revealed that the group getting both vitamin supplementation and exercise (CBD) had significantly higher osteoblast counts compared to the group receiving exercise alone (EXC) (Estimate = -11.52, SE = 5.22, p = 0.035). The CBD group did not exhibit any statistically significant differences when compared to both the control group and the group that only received vitamin supplementation (p > 0.05).
Table 6: Kruskal Wallis Test for Osteoclast and Cortex Thickness
|
Variable |
Chi square | P-value | Effect Size |
| Osteoclast count | 7.25 | 0.064 |
0.15 |
|
Cortex thickness |
1.97 | 0.579 |
-0.04 |
Table 7: Mann Whitney U test for osteoclast cell
|
|
U-Value | P-Value | Mean Rank Diff | Effect Size |
|
CON-EXC |
40.0 | 0.263 | 2.16 |
0.214 |
| CON-VIT | 38.0 | 0.430 | 1.65 |
0.297 |
|
CON-CBD |
39.0 | 0.036* |
4.14 |
0.581 |
|
EXC-VIT |
37.0 | 0.683 | -0.78 | 0.112 |
| EXC-CBD | 43.0 | 0.087 | 2.92 |
0.488 |
|
VIT-CBD |
45.5 | 0.060 | 3.55 |
0.447 |
(a) * significantly different (p<0.05)
The Shapiro-Wilk test revealed that the osteoclast counts data did not follow a normal distribution (p < 0.001), even though the variances were found to be uniform (p = 0.86). The number of osteoclasts was assessed using the Kruskal-Wallis test, which showed that there were no significant differences across the mice groups (F(3,28) = 2.222, p = 0.108). Table 7 shows further Mann-Whitney tests showed that the EXC group had significantly higher osteoclast counts than the CBD group (Estimate = 0.714, SE = 0.290, p = 0.020). There were no statistically significant disparities seen between the control group and the other groups, with a p-value greater than 0.05.
Table 8: Mann Whitney U Test for Cortex Thickness
|
U Value |
P Value | Mean Rank Diff |
Effect Size |
|
|
CON-VIT |
25.0 | 0.525 | -1.65 | 0.172 |
| CON-EXC | 35.0 | 0.751 | 0.89 |
0.0926 |
|
CON-CBD |
18.0 | 0.443 | -1.86 | 0.222 |
| VIT-EXC | 55.0 | 0.216 | 3.22 |
0.302 |
|
VIT-CBD |
33.0 | 0.916 | 0.38 | 0.0397 |
| EXC-CBD | 23.0 | 0.397 | -2.16 |
0.225 |
The Shapiro-Wilk test revealed that the data for cortical thickness did not follow a normal distribution (p = 0.000003), and Levene’s test suggested that the variances were not equal (p = 0.15). Analysis of femoral cortical thickness showed no significant variations across the groups (F(3,28) = 0.362, p = 0.781). Post hoc Mann-Whitney testing (Table 8) verified these findings, indicating that there were no substantial differences in cortical thickness across any of the groups (p > 0.05).
Discussion
Osteoblasts, osteoclasts, and osteocytes are essential cells in the bone microenvironment that engage in extracellular communication to collaborate and regulate bone remodelling. We can determine the precise state of the bone by studying these cells. Multiple studies have demonstrated that high-intensity physical exercise has a direct impact on bone metabolism. This impact is negative, as it leads to elevated levels of glucocorticoids and c-telopeptide, which in turn cause the death of osteocyte cells through a process called apoptosis.12,15
Oxidative stress, as defined by Sies (2017), is characterized by an imbalance in the equilibrium between prooxidants and antioxidants, with a bias towards the prooxidant side. Reactive oxygen species (ROS) are often associated with oxidative stress, which leads to severe consequences through the destruction of lipids, proteins, and DNA. Aerobic metabolism generates these reactive oxygen species (ROS), which include superoxide anion, hydrogen peroxide, and hydroxyl radicals. Each of these compounds has distinct chemical features that render them responsive to different biological targets.16,17
Reactive oxygen species (ROS) can disrupt normal bone remodelling processes by either being present in excessive amounts or being insufficient, which affects the growth and development of osteoclasts and osteoblasts. This ultimately leads to a reduction in bone density.18 Through the interaction between RANKL and its receptor, reactive oxygen species (ROS) are made, which causes osteoclasts to break down more bone. Furthermore, ROS suppresses osteoblastic cell development through the action of hydrogen peroxide. As a result, oxidative stress significantly impacts bone cell function by affecting both cell types and their intercellular communication.19
This study demonstrated that the high-intensity physical exercise treatment group had the fewest osteoblast and osteocyte cells (25.86 ± 4.667 and 26.86 ± 4.667, respectively). Significant differences in osteocyte count were observed between the group receiving only exercise compared to the group receiving combined treatment (p = 0.006) and the group receiving vitamin C and E supplementation only (p = 0.017). However, the exercise-only group and the combined treatment group showed a significant difference in osteoblast count (p = 0.035). As muscles contract, they produce free radicals, and prolonged and intense physical activity can cause oxidative damage to cellular structures8. Exercise recognizes the mitochondrial transport chain, polymorphonuclear leukocytes, and xanthine oxidase as important contributors to the generation of intracellular free radicals. The finding highlights the significance of oxidative stress in the body’s reactions to physical activity, as referenced by Ji.20
Experimental research by Biswajit Chaki showed that high-intensity exercise treatment elevated oxidative stress in both prepubertal and post pubertal men. The analysis of serum parameters, specifically the total activity of superoxide dismutase (SOD) and catalase (CAT), enzymes that protect cells from free radical damage, demonstrated this.18 In line with the idea and previous research, it has been discovered that the body’s defences may not be able to handle the production of free radicals or reactive oxidative chemicals. This can cause oxidative stress, which hurts the bone remodelling matrix made up of osteoblasts, osteoclasts, and osteocytes.21 An analysis of research papers by Emanuel. found that, in the long run, engaging in high levels of daily spontaneous physical activity is more beneficial for bone tissue than participating in high-intensity interval training programs. However, when combined with more intense physical activity, it can have negative effects on bone tissue.22,23 This state is a result of reactive oxygen species (ROS), which can directly increase the activity of osteoclasts by enhancing the interactions between RANKL and RANK. Additionally, ROS can affect the proteins P53 and foxO, leading to an accelerated death of osteoblasts.21
The group that did high-intensity exercise and was given Vitamins C and E had more osteoblast cells (37.86 ± 3.635) and osteocyte cells (54.7 ± 7.154) than the other groups. Additionally, the number of osteoclast cells, which indicates bone resorption, was the lowest in the CBD group (0.714 ± 0.184) and significantly different from the control group. An in vitro study examining the impact of Vitamins C and E on osteoblasts reveals that the injection of vitamin C (ascorbic acid) leads to an increase in osteoblast cells. We attribute this rise to enhanced protein synthesis of extracellular matrix collagen type I, osteonectin, and osteocalcin.9 A study, consistent with our findings, found that smokers who received Vitamin C and Vitamin E supplementation had a decreased chance ratio of hip fractures. The observation suggests that oxidative stress might play a role in the negative impact of smoking on bone health. Supplementing with Vitamin C and E could potentially provide protective advantages.24 Free radicals cause oxidative stress, which Vitamin E, a powerful antioxidant, reduces to protect bones. An experiment showed that treating osteoblasts with tocotrienol (1 μM) for 24 hours protected them from the harmful effects of hydrogen peroxide 25 in a model made by H2O2.25
In 2023, a recent study investigated the effects of vitamin C and E supplementation and high-intensity exercise on haematological variables. The study found that antioxidant supplementation with vitamins C and E offered physiological protection against exercise-induced haemolysis. An increase in red blood cell count and haemoglobin content, along with changes in white blood cell and platelet levels, demonstrated this protection. Additionally, the supplementation led to an improved inflammatory profile.26
Several influential aspects, such as the specific type of physical activity, the duration of therapy, and representative indicators of bone examined through cellular and molecular mechanisms, influence the disparity in the beneficial and detrimental impacts of high-intensity physical exercise, as shown in numerous studies. Theoretically, one of the reasons for apoptosis in the constituent cells of bone formation is inflammation. Reactive free radicals in the body can initiate inflammation by modifying lipids and proteins in living organisms, causing damage to tissues. The term used to describe this condition is reactive oxygen species (ROS).
This study has multiple drawbacks. Initially, the researcher relied on his expertise to establish the experiment’s dosage. Furthermore, it is critical to acknowledge that the experiment was specifically planned for a short period, possibly resulting in the oversight of any enduring consequences. Furthermore, the absence of measurements for MDA levels hindered our ability to appropriately evaluate oxidative stress levels. Further research with extended timeframes and direct data collection is necessary to confirm these results and gain a more thorough understanding of the impact of intense exercise and antioxidant supplements on bone health.
Conclusion
In conclusion, high-intensity exercise appears to increase bone resorption in mice, which could have a negative impact on bone health. On the other hand, taking antioxidant supplements containing vitamins C and E can stimulate the growth of bones and provide protection against the harmful effects of intense activity. The coalescence of vitamins C and E with vigorous exercise, demonstrates promising prospects for bone restructuring. Hence, it is imperative to provide those who partake in high-intensity exercise with antioxidant supplements, as they play a crucial role in safeguarding bone health. Further research is required to understand this mechanism and provide better solutions for maintaining bone health.
Acknowledgement
The authors would like to thank Universitas Airlangga for providing the opportunity to conduct research at Physiology and Medical Biochemistry, Universitas Airlangga, it is highly appreciated for allowing laboratory work. The authors are also very grateful to the Farma Veterinary Center (PUSVETMA) and the Anatomical Pathology department laboratory.
Funding Sources
This research is supported by Universitas Airlangga in the Faculty Excellence Research (PUF) research scheme managed by the internal research and community service institute of Universitas Airlangga.
Conflict of Interest
The authors have no conflict of interest to declare.
Data Availability Statement
This statement does not apply to this article
Ethics Statement
This research has been assessed and received permission from the Health Research Ethics Committee of the Faculty of Medicine, Universitas Airlangga, with certificate number 40/EC/KEPK/FKUA/2019.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials
Author Contributions
- Gadis Meinar: Conceptualization, Funding Acquisition, Idea and study design, Supervision, and Review.
- Julian Benedic: Writing, Visualization, Data analysis and interpretation, and Editing
- Yoga Akbar: Writing, Data analysis and interpretation, and Editing
- Lilik Herawati: Conceptualization, Idea and study design
- Irfiansyah Irwadi: Data collection and validation
- Silvia Maya: Data collection and writing.
References
- World Health Organization. Key facts. Water Conv 30 Years Impact Achiev Gr. 2022;(October):1-9. doi:10.18356/9789210018647c003
CrossRef - Hart NH, Nimphius S, Rantalainen T, Ireland A, Siafarikas A, Newton RU. Mechanical basis of bone strength: influence of bone material, bone structure and muscle action. J Musculoskelet Neuronal Interact. 2017;17(3):114-139.
- Carr AC, Maggini S. Vitamin C and immune function. Nutrients. 2017;9(11):1-25. doi:10.3390/nu9111211
CrossRef - Shivani Sahni, Marian T. Hannan, David Gagnon, Jeffrey Blumberg, L. Adrienne Cupples, Douglas P. Kiel and KLT. High Vitamin C Intake Is Associated with Lower 4-Year Bone Loss in Elderly Men. Bone. 2008;23(1):1-7. doi:10.1093/jn/138.10.1931
CrossRef - Lee PL, Lukman HI, Nazrun AS, Ima-Nirwana S, Norazlina M. Effects of vitamin E supplementation on bone metabolism in nicotine-treated rats. Singapore Med J. 2007;48(3):195-199.
- Rizvi S, Raza ST, Ahmed F, Ahmad A, Abbas S, Mahdi F. The role of Vitamin E in human health and some diseases. Sultan Qaboos Univ Med J. 2014;14(2):157-165.
- Gomez-Cabrera MC, Domenech E, Viña J. Moderate exercise is an antioxidant: Upregulation of antioxidant genes by training. Free Radic Biol Med. 2008;44(2):126-131. doi:10.1016/j.freeradbiomed.2007.02.001
CrossRef - Powers SK, Nelson WB, Hudson MB. Exercise-induced oxidative stress in humans: Cause and consequences. Free Radic Biol Med. 2011;51(5):942-950. doi:10.1016/j.freeradbiomed.2010.12.009
CrossRef - Urban K, Höhling HJ, Lüttenberg B, Szuwart T, Plate U. An in vitro study of osteoblast vitality influenced by the vitamins C and E. Head Face Med. 2012;8(1):1-10. doi:10.1186/1746-160X-8-25
CrossRef - Sarisözen B, Durak K, Dinçer G, Bilgen OF. The effects of vitamins E and C on fracture healing in rats. J Int Med Res. 2002;30(3):309-313. doi:10.1177/147323000203000312
CrossRef - D. R. Laurence and A. L. Bacharach. Evaluation of Drug Activities: Pharmacometrics. Vol 1.; 1964. doi:10.1021/jm00321a067
CrossRef - Sari GM, Soetjipto, Herawati L. Mechanism of Bone Metabolism Interruption Due to High Intensity Physical Exercise. Syst Rev Pharm. 2020;11(10):836-843. doi:10.31838/srp.2020.10.125
- Egan KP, Brennan TA, Pignolo RJ. Bone histomorphometry using free and commonly available software. Histopathology. 2012;61(6):1168-1173. doi:10.1111/j.1365-2559.2012.04333.x
CrossRef - Cerri PS. Osteoblasts engulf apoptotic bodies during alveolar bone formation in the rat maxilla. Anat Rec – Part A Discov Mol Cell Evol Biol. 2005;286(1):833-840. doi:10.1002/ar.a.20220
CrossRef - Mohr M, Helge E W, Petersen L F, Lindenskov A, Weihe P, Mortensen J, Jørgensen N R KP. Effects of soccer vs swim training on bone formation in sedentary middle-aged women. Eur J Appl Physiol. 2015;115(12):2671-2679. doi:10.1007/s00421-015-3231-8
- Helmut Sies, Carsten Berndt and DPJ. Oxidative Stress. Annual Review of Biochemistry. Annu Rev Biochem. 2017;86(1):715-748. doi:10.1146/annurev-biochem-061516-045037
CrossRef - Schieber M, Chandel NS. ROS function in redox signaling. Curr Biol. 2014;24(10):453-462. doi:10.1016/j.cub.2014.03.034.ROS
CrossRef - Biswajit Chaki, Sangita Pal SC& AB. High-intensity exercise-induced oxidative stress in sedentary pre-pubertal & post-pubertal boys: A comparative study. Indian J Med Res. 2019;(August):167-174. doi:10.4103/ijmr.IJMR
CrossRef - Wauquier F, Leotoing L, Coxam V, Guicheux J, Wittrant Y. Oxidative stress in bone remodelling and disease. Trends Mol Med. 2009;15(10):468-477. doi:10.1016/j.molmed.2009.08.004
CrossRef - Li li ji. Antioxidants and Oxidative Stress in Exercise (44453). Exp Biol Med. 2000;(44453):283-292. doi:10.1046/j.1525-1373.1999.d01-145.x
CrossRef - Ardies CM. Diet, Exercise,and Chronic Disease.; 2014. doi:10.4324/9781351022620-9
CrossRef - Polisel EEC, Beck WR, Scariot PPM, Pejon TMM, Gobatto CA, Manchado-Gobatto FB. Effects of high-intensity interval training in more or less active mice on biomechanical, biophysical and biochemical bone parameters. Sci Rep. 2021;11(1):1-11. doi:10.1038/s41598-021-85585-9
CrossRef - Manaye, Sara; Cheran, Kaaviya; Murthy, Chinmayee; Bornemann, Elisa A; Kamma, Hari Krishna; Alabbas, Mohammad; Elashahab, Mohammad; Abid, Naushad; Arcia Franchini AP. The Role of High-intensity and High-impact Exercises in Improving Bone Health in Postmenopausal Women: A Systematic Review. Cureus. 2023;15(2):1-8. doi:10.7759/cureus.34644
CrossRef - Holmberg L, Wolk A, Melhus H, Michae K. Smoking, Antioxidant Vitamins, and the Risk of Hip Fracture ¨. 1999;14(1). doi:https://doi.org/10.1359/jbmr.1999.14.1.129
CrossRef - Yongjie, Chen; Naichun, Yu; Daguo, Zhou; Zongguang, Li; Fengqing, Gong; Weijiang, Yi; Botao, Chen; Guangrong J. Vitamin E Inhibits Osteoclastogenesis in Protecting Osteoporosis. Vitam B Vitam E – Pleiotropic Nutr Benefits. Published online 2024:1-31. doi:10.5772/intechopen.112760
CrossRef - Sarkar S, Dey SK, Datta G, Bandyopadhyay A. Vitamin C and E supplementation and high intensity interval training induced changes in lipid profile and haematological variables of young males. Sport Med Heal Sci. 2023;5(2):137-145. doi:10.1016/j.smhs.2023.03.006
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






