
Introduction
Edible insects are becoming increasingly appreciated as an alternative to high-protein food, with consumers preferring processed insect products.1 Developing foods to persuade consumers to accept and eat more insects has become a major challenge, especially in regions where the consumption of whole insects lacks sensory appeal. When cricket powder is processed into hydrolyzed protein, it has a higher water solubility and can be used in a wider range of foods. Hydrolyzed protein is produced using different methods to hydrolyze proteins into short peptides and free amino acids. The structures, characteristics, and chemistry of proteins are all impacted by degradation. Acids and alkalis are commonly used as a quick and inexpensive method to hydrolyze the degradation of protein peptide bonds. However, chemicals used during production must be removed before use and may leave residue. Enzymatic analysis can improve the properties of the protein by using mild conditions and controlling the level of digestibility. The rate of decomposition using this method is also high compared to using acids and alkalis. Enzymatic hydrolysis of insect protein is an alternative method for improving the properties of its protein hydrolysate. Enzymatic hydrolysis can enhance protein extraction yield, increase the degree of hydrolysis (DH), and liberate free amino acids. Hydrolyzing Tenebrio molitor protein demonstrated improved yield, higher DH, and greater release of free amino acids.2 Additionally, treatment with alcalase and flavoenzyme further enriched the amino acid profile and enhanced the product’s scavenging potential.3 Alcalase hydrolysis was also used to improve the technological and functional properties of protein flour produced from migrating locusts (Locusta migratoria L.).4 Therefore, enzymatic hydrolysis is an alternative process for high protein edible insects, improving the functional and health-promoting properties of the resulting protein hydrolysates
Crickets are a popular insect with protein quality and digestibility comparable to chicken, pork, and beef.5 In particular field cricket (Gryllus bimaculatus) has a high protein concentration of approximately 60 to 70% dry weight, which is comparable to traditional animal proteins.6 The nutritional value and practical uses of cricket components as food ingredients have recently attracted increased attention but few studies have addressed the functional characteristics of proteins from different insects. Insect hydrolysate effectiveness is contingent on protein characteristics such as high solubility and gel, foam, or emulsion formation stability. Protein hydrolysates can be applied across domains, including functional attributes like foams and emulsifiers. Using protein hydrolysates from edible crickets as an alternative protein source is a desirable choice in the context of global food security and nutrition. However, there is no information available about hydrolyzing edible cricket proteins with commercial enzymes such as alcalase, as well as the biological properties of protein hydrolysate produced. The purpose of this work was to investigate the effects of alcalase hydrolysis time on the physicochemical and functional properties of protein hydrolysates extracted from G. bimaculatus cricket powder. The results were compared to an enzyme-free protein hydrolysate.
Materials and Methods
Chemicals
Unless otherwise specified, all chemicals used were of reagent grade and sourced from Sigma Aldrich (St. Louis, MO, USA) and Thermo Fisher Scientific (Waltham, MA, USA). Alcalase® (a protease derived from Bacillus licheniformis, P2.4 U/g) was obtained from Sigma Aldrich. Angiotensin-converting enzyme (ACE) from rabbit lung and the substrate hippuryl-L-histidyl-L-leucine (HHL) were procured from Dinojo, Japan.
Raw materials
Frozen Gillus bimaculatus were purchased from a local cricket farm in Maha Sarakham Province, Thailand. The crickets were cultivated under good agricultural practice (GAP) guidelines and harvested as adult crickets at 42-45 days old.7
Cricket powder preparation
After thawing and blanching the frozen crickets in a 1:10 (w/w) ratio for 10 min at 80°C, they were cooled in water at 10±1°C. A tray dryer was used immediately to dehydrate them. Three kilograms of crickets were divided equally among the drying trays and dried at 80°C (Memmert, OLM-500, Germany) until the water activity (aw) was less than 0.6 and the moisture content was 10±1% (wet basis). Using the Kjeldahl method, the protein content of the finely ground powdered dried crickets was found to be 62.15%.8
Cricket protein hydrolysate preparation
Cricket protein hydrolysate was produced with minor alterations following previous procudure.9 The cricket powder (14.0 g) was dissolved in 70 mL of 0.2 M phosphate buffer (RCI Labscan, USA) at pH 8, and then added to alcalase enzyme (Sigma, USA) at a concentration of 4% by weight of protein in the powder (60.31%). Hydrolysis was carried out in a continuously shaking water bath (Lauda, Germany) at 50°C for 180 min, with samples collected at 0, 20, 40, 60, 120, and 180 min before the digestion was ended by heating to 95°C for 10 min. The samples were centrifuged for 30 min at 4,000 rpm using a Hettich centrifuge (UNIVERSAL 320). The clear solution (supernatant) was frozen at -30°C and then freeze-dried to obtain cricket protein hydrolysate (CPH). All experiments were conducted in triplicate.
Determination of degree of hydrolysis
The DH was measured using the hydrolysis rate of o-phthaldialdehyde (OPA).10 The protein hydrolysate powder was dissolved in distilled water (1 mg/mL) and 25 µL of protein hydrolysate solution was added to 188 µL of OPA reagent (Sigma, USA) and mixed at 25°C for 2 min. The absorbance of the mixture solution was recorded using a microplate reader (Spectrostar Nano, Germany) at 340 nm. The rate of degradation was computed using the equation:
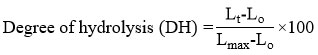
where DH (%) represents the degradation rate percentage,
Lt = the concentration of amino acids presents at different digestion times of protein hydrolysate.
Lo = the concentration of amino acids presents at the start of the mixture of cricket powder and phosphate buffer.
Lmax was determined by measuring the total quantity of amino acids after the cricket powder was completely hydrolyzed with 12 N HCl for 24 hours (1:1) at 90°C.
Amino acid content analysis
Each 25 mL tube was filled with a 1.0 mL aliquot of CPH solution (1% in water), followed by the addition of 0.8 mL of deionized water, 1.0 mL of phosphoric acid buffer solution (pH 6.0), and 1.0 mL of 2% ninhydrin solution. After 15 min of reaction at 90°C, the mixtures’ volume was adjusted to 25 mL using deionized water. After measuring the solutions’ absorbance at 570 nm, the free amino acid (FAA) content was computed using leucine-prepared standard curves.
Proximate composition of cricket powder
The proximate compositions of CPH were determined using the AOAC official method (2000).8
Protein content of liquid cricket protein hydrolysate
The protein contents of cricket hydrolysate samples collected at different hydrolysis times were determined by the Lowry method.11
Protein solubility (%)
The CPH solubility was assessed using the modified method.9 200 mg of CPH and the control samples were dissolved in 20 mL of pH 7.0 buffer and agitated with a magnetic stirrer for 30 min at room temperature. After 15 min at 4°C, the mixture was centrifuged at 10,000 ×g. The protein content of the supernatant and the total protein content of each sample were determined using the Lowry method. Using the following formula, protein solubility was calculated and expressed as a percentage.

Foaming ability and foam stability
The aeration method was utilized to assess foaming ability (FA). A 25 mL of phosphate buffer (pH 7.0) was used to dissolve a 4 mL aliquot of CPH. A stir bar was used to equilibrate the mixture for 10 min at room temperature. A homogenizer set to 12,000 rpm for 2 min was then used to aerate the mixture and produce bubbles (IKA, USA).4 The volume of the foam produced was measured by pouring it into a 25 mL measuring cylinder. The proportion of foam that remained after 30 min was used to compute the foaming ability and foaming stability (FS) as follows:

Emulsifying activity
The emulsifying activity (EA) was evaluated using a previously reported method with minor modifications.4 Briefly, Protein solutions (4%) were made by mixing CPH and the control sample with phosphate buffer pH 7.0. The samples were centrifuged at 3,388×g for 15 min after being allowed to equilibrate for two hours at room temperature. The supernatant was homogenized for one min at 12,000 rpm after being mixed with pure sunflower oil (1:1 v/v). Using scaled tubes, 15 mL aliquots of the emulsion were centrifuged at 3,388 ×g for 15 min at room temperature. The height of the overall height of the solution (HS) and the height of the resulting emulsified layer (HEL) were used to calculate the EA (Eq. 4) as follows:
![]()
Determination of antioxidant activity
The CPH samples were treated with and without enzyme hydrolysis at various interval periods.7 A 0.3 g portion of each sample was extracted in 25 mL of 80% ethanol at 37°C for 18 hours using a shaker (Mettler Toledo, AG8603 Schwerzenbach). The mixture was filtered via filter paper after being centrifuged for 30 min at 4,000 ×g. The total phenolic content (TPC) and antioxidant activity of the extract samples were assessed.
Diphenyl–2–picrylhydrazyl (DPPH) assay
DPPH radical scavenging activity was assessed.12 The results were compared with a reference standard (gallic acid) and expressed as mg GAE/g dry basis of a sample.
2,2-azino-bis-3-ethylbenzothiazoline-6-sulphonic acid (ABTS) radical scavenging assay
The ABTS radical cation decolorization assay was used to measure the free radical scavenging activities of CPH and the control.13 The results were reported in mg GAE/g dry basis of a sample.
Ferric reducing antioxidant power (FRAP)
The ability of the CPH and control samples to change Fe3+ -TPTZ into blue-colored Fe2+-TPTZ was used to calculate their reducing power.14 The results were expressed in µM FeSO4/g dry basis of material, and the absorbance was recorded at 539 nm.
Total phenolic content (TPC) measurement
The TPC of the extracts was evaluated using the Folin-Ciocalteu reagent.15 At 765 nm, the absorbance was determined with a spectrophotometer (CE 2041, England). Gallic acid was used to create the standard curve, and the results were reported as GAE/g dry basis of the sample.
Determination of angiotensin-converting enzyme
An ACE Kit-WST (Dojindo Laboratories Technologies Inc., Japan) was used to evaluate the angiotensin-converting enzyme (ACE).16 Enzyme B was dissolved in 2 mL of distilled water to create the enzyme working solution, which was then added to enzyme A in 1.5 mL. Enzyme C and the coenzyme were dissolved in 3 mL of purified water and thoroughly mixed to create the indicator working solution. After adding 20 µL of each of the sample extract, substrate buffer, and enzyme working solution to a 96-well plate, the plates were incubated for one hour at 37°C (model IPS 260/750 Memmert: Germany) before 200 mL of the indicator working solution was added. A microplate reader was used to measure the sample mixture’s absorbance at 450 nm after it had been left in the dark at room temperature for 10 min. The following formula was then used to determine the ACE inhibition ability:
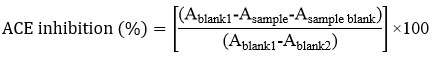
Statistical analysis
The results were presented as mean and standard deviation values for triplicate experiments. A one-way analysis of variance (ANOVA) was used to compare data from G. bimaculatus protein hydrolysis with and without alcalase at different times (0, 20, 40, 60, 120, and 180 minutes). Significant differences between treatments were identified using Duncan’s multiple range test. The threshold for statistical significance was established at p<0.05. All experiments were completed in triplicate.
Results
Effect of hydrolysis time on yields
Before hydrolysis, G. bimaculatus powder contained 62.15% protein and 20.22% fat. Table 1 displays the yields of CPH treated with and without alcalase hydrolysis (control) obtained from different hydrolysis times. The CPH yields increased significantly (p<0.05) when increasing the enzymatic hydrolysis duration, but no significant difference was recorded for yields in crickets without alcalase hydrolysis. Crickets hydrolyzed with alcalase for 120 and 180 min produced the highest yields of CPH (37.98% and 39.17%, respectively) while yields of crickets digested without adding enzyme varied from 23.35 to 25.05%.
Effect of hydrolysis time on DH
Alcalase hydrolysis time significantly affected the DH of G. bimaculatus crickets (p<0.05) (Table 1). Longer enzyme hydrolysis time yielded peptides with higher DH. Crickets hydrolyzed with alcalase for 120 and 180 min showed significantly (p<0.05) higher DH values at 84.78% and 84.42%, respectively whereas lower DH values were recorded in crickets without alcalase hydrolysis (control) ranging from 13.62 to 27.46%.
Effect of hydrolysis time on protein and free amino acid content
Table 1 shows the change in protein and free amino acid (FAA) contents of CPH obtained from each hydrolysis time during 180 min13.62 of hydrolysis. The protein and FAA contents in CPH gradually increased at longer alcalase hydrolysis times, whereas changes in the control samples were not significantly different.
Table 1: CPH yield, hydrolysis degree, protein, amino acids, and phenolic compounds with/without alcalase at different times
|
Treatment |
Time (min) | Yield (%) | DH (%) | Protein (mg/mL) | Free amino acid (mg/g) |
| Control | 0 | 23.85±0.04 e | 13.62±0.02e | 10.14±0.67e |
1.63±0.04f |
|
20 |
25.05±0.02e | 16.00±0.03d | 10.26±0.28e | 1.71±0.06e | |
| 40 | 23.35±0.04e | 18.69±0.02c | 10.96±0.67e |
1.91±0.01d |
|
| 60 | 24.16±0.09e | 19.35±0.01b | 11.01±1.38e | 1.97±0.01d | |
| 120 | 23.62±0.01e | 21.39±0.01c | 10.74±0.25e |
2.07±0.02cd |
|
| 180 | 24.43±0.04e | 27.46±0.02a | 10.85±0.40e | 2.11±0.07c | |
| Alcalase | 0 | 24.15±0.04e | 48.91±1.17e | 12.38±0.30e |
1.72±0.06f |
| 20 | 32.41±0.08d | 65.07±1.28d | 22.81±0.15d | 2.19±0.01c | |
| 40 | 33.97±0.14c | 75.50±0.82c | 25.06±0.46c |
3.32±0.03b |
|
| 60 | 36.25±0.05b | 79.81±1.17b | 29.96±1.88b | 4.73±0.06a | |
| 120 | 37.98±0.21ab | 84.78±1.35a | 31.12±0.64a |
4.88±0.04a |
|
| 180 | 39.17±0.19a | 84.42±1.21a | 31.67±1.44a |
4.94±0.06a |
Values are the means ± SD of dry weight samples taken in triplicate.
a-f significant differences are shown by different superscripts within a column (p<0.05).
Effect of hydrolysis time on TPC
No significant impact (p<0.05) on TPC was determined at different hydrolysis times. TPC in the control samples treated without alcalase was comparable to values in CPH produced by an enzymatic process (Figure 1).
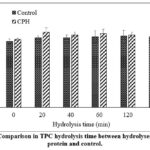 |
Figure 1: Comparison in TPC hydrolysis time between hydrolyses of cricket protein and control. |
Effect of hydrolysis time on protein solubility
Protein solubility at pH 7 gradually increased at longer hydrolysis times (Table 2). The highest protein solubility at 86.73% was recorded after 120 min, significantly higher than the control at the same hydrolysis time.
Effect of hydrolysis time on foaming properties
Foaming ability varied from 96.7 to 100% (Table 2), with the highest value recorded for alcalase CPH digestion at 60 min. Foaming stability ranged from 80.0 to 86.9% and was significantly higher than the control.
Effect of hydrolysis time on emulsification activity
The emulsification activity of CPH is shown in Table 2. EA values of all treatments were not significantly different and ranged from 49.5 to 52.4%.
Table 2: Protein solubility, foaming properties, and emulsion activity of CPH with/without alcalase at different times
|
Treatment |
Time (min) | Protein solubility (%) | Foaming properties | Emulsion activity (%) ns | |
| FA (%) | FS (%) | ||||
| Control | 0 | 21.25±1.2g | 44.0±1.4e | 40.4±1.5e |
51.6±0.9 |
|
20 |
23.16±1.7f | 44.0±2.8c | 56.0±1.0cd | 52.4±2.1 | |
| 40 | 23.67±2.8f | 42.5±0.7d | 55.5±0.7cd |
51.6±2.1 |
|
|
60 |
25.12±1.2f | 40.3±1.4f | 56.7±1.0e | 51.4±0.6 | |
| 120 | 32.55±1.7e | 42.6±1.1b | 60.2±1.4c |
51.1±1.3 |
|
|
180 |
33.14±1.7e | 40.2±1.2d | 52.0±0.7d | 51.6±0.9 | |
| Alcalase | 0 | 29.78±1.08e | 96.7±1.8 | 80.0±0.9b |
50.9±0.7 |
|
20 |
45.97±0.9d | 98.4±2.6 | 82.1±3.2b | 49.5±2.2 | |
| 40 | 66.51±1.5c | 99.0±1.4 | 84.7±2.1ab |
51.4±0.7 |
|
|
60 |
76.16±3.3b | 100.0±3.1 | 86.4±1.7a | 52.0±1.0 | |
| 120 | 86.73±1.1a | 99.1±2.0 | 86.9±2.9a |
51.2±1.4 |
|
|
180 |
85.34±2.8a | 98.3±2.5 | 86.1±2.7a |
51.1±1.3 |
|
Values are means ±SD of triplicate samples (dry weight).
a-f Different superscripts within a column indicate significant differences (p<0.05).
FA = foaming ability; FS = foaming stability.
Effect of hydrolysis time on antioxidant activity
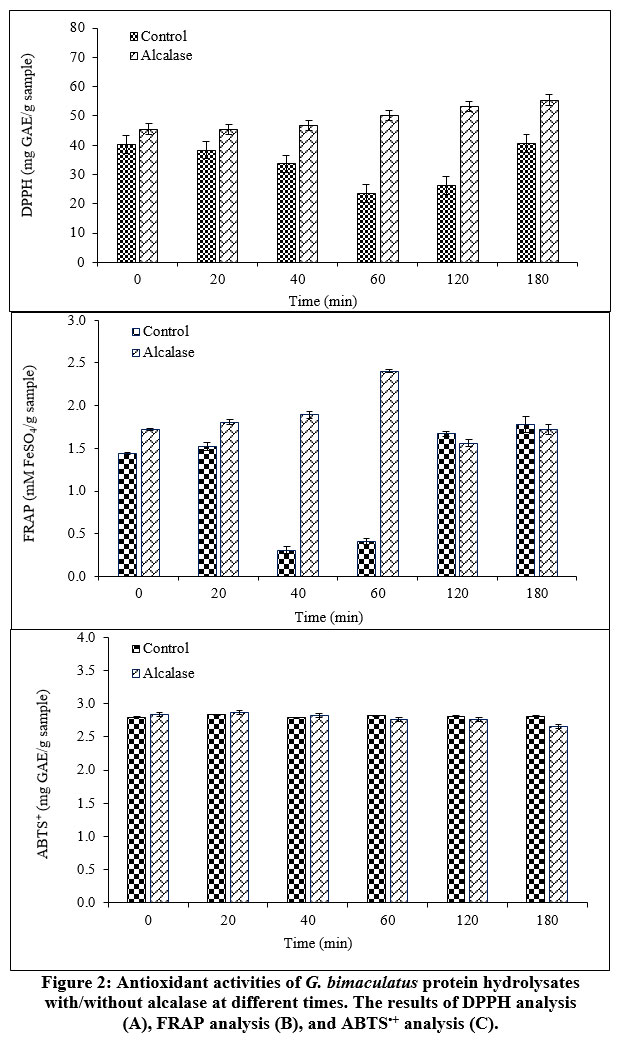
Figure 2 shows the antioxidant activities of CPH treated with and without alcalase hydrolysis at various times. Hydrolysis time significantly (p<0.05) impacted CPH DPPH and FRAP values but no significant differences in ABTS•+ values were found. The DPPH radical scavenging activity increased as alcalase hydrolysis time increased. CPH treated with alcalase gave the highest DPPH values at 60, 90, and 180 min (50.23, 53.25, and 55.43 mg GAE/g, respectively) while the control recorded the lowest value at 60 min (23.49 mg GAE/g). The FRAP values gradually increased with increasing alcalase hydrolysis times from 0 to 60 min, and then decreased after 120 and 180 min. The CPH treated with alcalase at 60 min had the highest FRAP value (2.40 mM FeSO4/g), whereas CPH without the enzyme at 30 min had a low FRAP value (0.31 mM FeSO4/g).
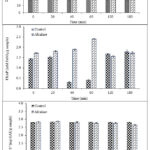 |
Figure 2: Antioxidant activities of G. bimaculatus protein hydrolysates with/without alcalase at different times. The results of DPPH analysis (A), FRAP analysis (B), and ABTS•+ analysis (C). |
Effect of hydrolysis time on ACE inhibition
The inhibition activity of ACE is one of the most common treatments for hypertension. Bioactive peptides derived from protein-rich foods are studied for their ability to reduce blood pressure by inhibiting ACE. Figure 3 shows that the ACE inhibition of CPH treated with and without alcalase at different times increased with increasing alcalase hydrolysis time. CPH treated with alcalase for 120 and 180 min showed the highest ACE inhibition values at 69.12 and 69.23%.
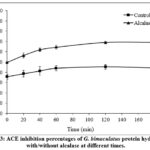 |
Figure 3: ACE inhibition percentages of G. bimaculatus protein hydrolyses with/without alcalase at different times. |
Proximate composition of CPH
The CPH obtained from 120 min digestion time was chosen for proximate composition analysis. The crude protein and ash contents were higher than the control, while moisture and fat contents were lower. The crude protein content of CPH was 64.89%, and higher than the control (62.15%) (Figure 4).
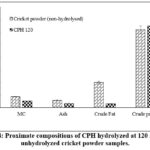 |
Figure 4: Proximate compositions of CPH hydrolyzed at 120 min and unhydrolyzed cricket powder samples. |
Discussion
The DH is an indicator of peptide bond cleavage, with structured proteins degrading into smaller peptides.4 Hydrolysis time significantly impacted the yield and DH. An increase in hydrolysis time enhanced the DH values of the cricket samples, resulting in a higher yield of CPH. The yield and DH of the control were lower than CPH because of no enzyme activity. Blanching as a pre-treatment and tray drying was used to inactivate the existing enzymes in G. bimaculatus cricket powder and denature its protein. Heating and water hydrolyzed the cricket powder, with increased yield and DH at 180 min of digestion. A heated cricket solution without enzyme addition has low DH (5.2%) because of protein denaturation and enzyme inactivation.17 Cricket samples hydrolyzed with low enzyme concentration (0.5%) had lower DH, while samples treated with 3% enzyme concentration revealed high DH after 60 minutes.9 Higher enzyme/substrate concentrations and longer hydrolysis times resulted in higher DH, while the control sample showed a low DH of 4.5%.18 Similarly, hydrolysis of Gryllodes sigillatus proteins with 3% alcalase produced the highest DH value of 52%.10 This study recorded the highest cricket proteins with alcalase hydrolysis after 120 minutes, indicating substrate active site enzyme saturation which prohibited further hydrolysis. However, DH values are impacted by several variables such as pH, temperature, hydrolysis time, and enzyme type and concentration. The functional and biological features of CPH products vary depending on the size, shape, amount of released peptides, and amino acid makeup.19 Each increase in hydrolysis time gave higher protein and FFA contents compared to the control. This finding concurred with earlier research which reported that amino acid content varied.10 In comparison to the control, some amino acid concentrations rose while others fell (for instance, Tyr and Arg levels were higher than the control). The FAA content (Val, Ile, Phe, Trp, Leu, and Tyr) that influenced taste was also improved by hydrolysis, especially tyrosine, which adds bitter or unpleasant flavors.20 According to the research, utilizing protein hydrolysates in foods may cause issues with bitter peptide formation.21 Foam development is controlled by molecular mobility, diffusion, and reorganization at the air-water interface. To be effective, foaming proteins must be able to quickly travel to the air-water contact, unfold, and rearrange at that interface, resulting in higher concentrations of amino acids during digestion. Amphiphilic proteins are also necessary for good surface characteristics. Different foam generation values are produced by the proteins and amino acid types found in each species of cricket. Hydrophilic amino acids are significant components of G. bimaculatus.7 At the air-water contact, globular proteins showed reduced ability to unfold which diminished form foaming capacity.
Hydrolysis had no noticeable effect on the emulsion’s capacity. CPH was found to be associated with significant enzymatic hydrolyzed protein and a reduction in emulsion activity.22 Although greater interfacial activity was thought to be responsible for CPHs improved foaming and emulsion capabilities, hydrolysis also decreased the molecular weight of the polypeptides, and polypeptide size affected the interfacial layers’ strength and network formation. Over time, hydrolysis weakens emulsions and foams’ ability to stay stable. Although hydrolyzed cricket proteins showed good foaming properties in this experiment, there was no discernible effect on emulsion ability. Bread and other bakery goods have shown promise in using CPH as a foaming agent alternative. No apparent change in ABTS•+ was observed in CPH’s antioxidant activity. DPPH and other peroxyl radicals are actively interacting with hydrophobic peptides, although ABTS•+ is more sensitive to hydrophilic peptides.19 The antioxidant activity of peptides with low molecular weight increases because they expose more amino acids to interactions with free radicals, while enzymatic hydrolysis reactions break down large molecular weight peptides into smaller hydrophobic di- or tri- peptides with higher antioxidant capacity, resulting in higher antioxidant activity.23 In CHP produced by enzymatic hydrolysis, some peptides contained amino acids (e.g., phenylalanine, histidine) associated with electron transfer capacity, resulting in increased antioxidant FRAP activity values. Protein hydrolysates have high reduced potential due to the availability of hydrogen protons and electrons generated by peptide bond cleavage.24 As reported in our previous study, G. bimaculatus cricket powder contains eight essential amino acids, six non-essential amino acids, and hydrophobic amino acids.7 Protein hydrolysates high in hydrophobic amino acids are linked to the ability to scavenge or inhibit free radicals.25
His, Phe, Tyr, Trp, or Met undergo physical or chemical one-electron oxidation that inhibits amino acid formation.26 Antioxidant substances including thiol groups reduce the sensitively of the FRAP technique.27 Protein hydrolysates show antioxidant activity through lipid peroxidation, electron transfer, oxidative interactions with oxygen-containing chemicals, inhibition of free radicals, and metal ionization.28 G. bimaculatus crickets are composed of amino acids and their protein hydrolysates show potential as bioactive substances with antioxidant activities.
Peptides derived from CPH hydrolyzed with alcalase exhibit strong in vitro ACE inhibition, particularly those with a higher DH. As a result, alcalase-treated CPH samples demonstrated superior ACE inhibition compared to the control sample. The findings agreed with those of 18 Hall et al. who discovered that ACE inhibition in CPH increased in tandem with DH. The C-terminal amino acids with hydrophobic aromatic amino acid residues (Tyr, Phe, and Trp) or hydrophobic branched side chains (Val, Leu, and Ile) have an impact on the ACE inhibitory action.29 These amino acids are usually associated with ACE-inhibitory activity and were found at the highest concentrations when hydrolyzed with 1.5% enzyme for 90 minutes.9,30 The CPH obtained contained higher protein content than non-hydrolyzed crickets. The alcalase-hydrolyzed samples had higher protein content than the control.9
Conclusion
The alcalase treatment increased the biological activity of CPH from G. bimaculatus. Higher alcalase hydrolysis duration resulted in considerably higher CPH yields and DH. CPH treated with the enzyme for 120 and 180 minutes yielded the highest CPH yields (37.98 and 39.17 g/100 g, respectively) and the highest DH levels (84.78% and 84.42%). These durations also resulted in the greatest antioxidant activity (DPPH) and ACE inhibition activity when compared to the control (untreated CPH). The findings demonstrate that alcalase treatment is an effective technique for producing cricket-derived protein hydrolysates with enhanced biological activity. CPH showed promise for utilization in food formulations, with high foaming ability and foam stability. The development is driven by the need for sustainable food production methods to meet global protein demands while addressing sustainability concerns.
Acknowledgement
The author would like to thank, Department of Food Technology and Nutrition and Laboratory Equipment Center, Mahasarakham University, Thailand for their guidance and support in completing this article.
Funding Sources
This research project was financially supported by Thailand Science Research and Innovation (TSRI).
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal subjects, or any materials that require ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to reproduce materials from other sources
Not applicable.
Clinical Trial Registration
This research does not involve any clinical trials.
Author Contributions
- Manatchaya Sangsri-In: Conceptualization, Methodology, Data analysis.
- Anuchita Moongngarm: Conceptualization, Methodology, Data analysis, Writing-review & editing, Supervision.
- Sirirat Deeseenthum: Fund acquisition, Methodology, Supervision.
- Vijitra Luang-In: Methodology, Supervision.
- Luchai Butkhup: Methodology, Supervision.
- Isaraporn Somboonwatthanakul: Methodology, Supervision.
- Suthathip Phankhlong: Methodology, Data analysis.
- Kedkasorn Yachai: Methodology, Data analysis.
- Yonlada Khawla: Methodology, Data analysis.
- Saranya Pimkoksoong: Methodology, Data analysis.
- Treechada Utaida: Methodology, Data analysis.
- Patiwit Loypimai: Data analysis, Writing-review & editing.
References
- Hartmann C., Siegrist M. Insects as food: Perception and acceptance. Ernährungs Umschau. 2017; 64(3): 44-50. doi:10.4455/EU.2017.010.
- Leni G., Soetemans L., Jacobs J., Depraetere S., Gianotten N., Bastiaens L., Caligiani A., Sforza S. Protein hydrolysates from Alphitobius diaperinus and Hermetia illucens larvae treated with commercial proteases. J Insects Food Feed. 2020; 6(4): 393-404. org/10.3920/JIFF2019.0037.
CrossRef - Tang Y., Debnath T., Choi E.J., Kim Y.W., Ryu J.P., Jang S., Chung S.U., Choi Y.J., Kim E.K. Changes in the amino acid profiles and free radical scavenging activities of Tenebrio molitor larvae following enzymatic hydrolysis. PloS one. 2018; 13(5): e0196218. doi.org/10.1371/journal.pone.0196218.
CrossRef - Purschke B., Meinlschmidt P., Horn C., Rieder O., Jäger, H. Improvement of techno-functional properties of edible insect protein from migratory locust by enzymatic hydrolysis. Eur Food Res Technol. 2018; 244(6): 999-1013. org/10.1007/s00217-017-3017-9.
CrossRef - Murugu D.K., Onyango A.N., Ndiritu A.K., Osuga I.M., Xavier C., Nakimbugwe D., Tanga C.M. From farm to fork: Crickets as alternative source of protein, minerals, and vitamins. Front Nutr. 2021; 10(8): 704002. doi:10.3389/fnut.2021.704002.
CrossRef - Udomsil N., Imsoonthornruksa S., Gosalawit C., Ketudat-Cairns M. Nutritional values, and functional properties of house cricket (Acheta domesticus) and field cricket (Gryllus bimaculatus). Food Sci Technol Res. 2019; 25(4): 597-605. doi.org/10.3136/fstr.25.597.
CrossRef - Loypimai P., Moontree T., Pranil T., Moongngarm A. A comparative study of nutritional components of Gryllus bimaculatusand Acheta domesticus cricket powder prepared using different drying methods. Food Measure.2024;18(5): 3974-3983.org/10.1007/s11694-024-02469-y.
CrossRef - Official Methods of Analysis. 17th ed. Washington, D.C.: Association of Official Analytical Chemical. 2000.
- Hall F.G., Jones O.G., O’Haire M.E., Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017; 224(6): 414-422. org/10.1016/j.foodchem.2016.11.138.
CrossRef - Irshad I., Kanekanian A., Peters A., Masud T. Antioxidant activity of bioactive peptides derived from bovine casein hydrolysate fractions. J Food Technol. 2015; 52(1): 231-239. doi.org/10.1007/s13197-012-0920-8.
CrossRef - Hartree E.F. Determination of protein: A modification of the Lowry method that gives a linear photometric response. Anal Biochem. 1972; 48(2): 422-427. doi.org/10.1016/0003-2697(72)90094-2.
CrossRef - Dasgupta N, De, B. Antioxidant activity of Piper betle L. leaf extract in vitro. Food Chem. 2004; 88(2): 219-224. org/10.1016/j.foodchem.2004.1001.1036.
CrossRef - Moongngarm A., Sriharboot N., Loypimai P., Moontree T. Ohmic heating-assisted water extraction of steviol glycosides and phytochemicals from Stevia rebaudiana LWT. 2022; 154(1): 112798. doi.org/10.1016/j.lwt.2021.112798.
CrossRef - Benzie I.F., Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: the FRAP assay. Anal Biochem 1996; 239(1): 70-76.
CrossRef - Iqbal S., Bhanger M.I., Anwar F. Antioxidant properties and components of some commercially available varieties of rice bran in Pakistan. Food Chem. 2005; 93(2): 265-272. org/10.1016/j.foodchem.2004.1009.1024.
CrossRef - Martínez-Alvarez O., Batista I., Ramos, C., Montero P. Enhancement of ACE and prolyl oligopeptidase inhibitory potency of protein hydrolysates from sardine and tuna by-products by simulated gastrointestinal digestion. Food Funct.2016; 7(4): 2066-2073. doi.org/10.1039/c5fo01603g.
CrossRef - Chi E.Y., Krishnan S., Randolph, T.W., Carpenter, J.F. Physical stability of proteins in aqueous solution: Mechanism and driving forces in nonnative protein aggregation. Pharm Res. 2003; 20(9): 1325-1336. doi.org/10.1023/A:1025771421906.
CrossRef - Hall F., Johnson P.E., Liceaga A. Effect of enzymatic hydrolysis on bioactive properties and allergenicity of cricket (Gryllodes sigillatus) Food Chem. 2018; 262(10): 39-47. doi.org/10.1016/j.foodchem.2018.04.058.
CrossRef - Chalamaiah M., Dinesh Kumar B., Hemalatha R., Jyothirmayi T. Fish protein hydrolysates: proximate composition, amino acid composition, antioxidant activities and applications: a review. Food Chem. 2012; 135(4): 3020-3038. org/10.1016/j.foodchem.2012.06.100.
CrossRef - Pedersen B. Removing bitterness from protein hydrolysates. Food Technol. 1994; 48(10): 96-98.
- Clemente A. Enzymatic protein hydrolysates in human nutrition. Trends Food Sci Technol. 2000; 11(7): 254-262. doi: 10.1016/S0924-2244(01)00007-3.
CrossRef - Chen C., Chi Y.J., Zhao M.Y., Xu W. Influence of degree of hydrolysis on functional properties, antioxidant and ACE inhibitory activities of egg white protein hydrolysate. Food Sci Biotechnol. 2012; 21(2): 27-34. org/10.1007/s10068-012-0004-6.
CrossRef - Liu Y., Wan S., Liu J.U.N., Zou Y., Liao S. Antioxidant activity and stability study of peptides from enzymatically hydrolyzed male silkmoth. J Food Process Preserv. 2017; 41(1): e13081. org/10.1111/jfpp.13081.
CrossRef - Ketnawa S., Liceaga A.M. Effect of microwave treatments on antioxidant activity and antigenicity of fish frame protein hydrolysates. Food Bioprocess Techno. 2017; 10(3): 582-591. org/10.1007/s11947-016-1841-8.
CrossRef - Saadi S., Saari N., Anwar F., Hamid A.A., Ghazali H.M. Recent advances in food biopeptides: production, biological functionalities and therapeutic applications. Biotechnol Adv. 2015; 33(1): 80-116. doi: 10.1016/j.biotechadv.2014.12.003.
CrossRef - Elias R.J., Kellerby S.S., Decker E.A. Antioxidant activity of proteins and peptides. Crit Rev Food Sci Nutr. 2008; 48(5): 430e441. org/10.1080/10408390701425615.
CrossRef - Liang N., Kitts D. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. 2014; 19(11): 19180e19208. doi.org/10.3390/molecules191119180.
CrossRef - Chen H-M., Muramoto K., Yamauchi F., Nokihara K. Antioxidant activity of designed peptides based on the antioxidative peptide isolated from digests of a soybean protein. J Agric Food Chem. 1996; 44(9): 2619-2623. org/10.1021/jf950833m.
CrossRef - Dai C., Ma H., Luo L., Yin, X. Angiotensin I-converting enzyme (ACE) inhibitory peptide derived from Tenebrio molitor (L.) larva protein hydrolysate. Eur Food Res Technol. 2013; 236(4): 681-689. org/10.1007/s00217-013-1923-z.
CrossRef - Megías C., del Mar Yust M., Pedroche J., Lquari H., Girón-Calle J., Alaiz M., Millán F., Vioque, J. Purification of an ACE inhibitory peptide after hydrolysis of sunflower (Helianthus annuus) protein isolates. J Agric Food Chem. 2004; 52(7): 1928-1932. http://dx.doi.org/10.1021/jf034707r. PMid:15053531.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.







{kind=link}