
Introduction
Humans utilize various plants as food sources at different stages of their fruit development, including peppers, cucumbers, and dates. The characteristics of these fruits, such as color, taste, aroma, and firmness, vary throughout their development stages. Numerous studies have indicated that fruits display both qualitative and quantitative differences in their chemical composition during ripening.1,2 These variations are influenced by genetic factors, agricultural practices, climatic conditions, and storage methods.3–5 The date palm, classified under the family Arecaceae (Palmaceae), is commonly cultivated in regions such as North Africa, the Middle East, and South Asia. 6 The fruit of the date palm, known as dates, has served as an important food source and a means of wealth in the deserts of North Africa and the Middle East for centuries.7 Dates are an excellent source of carbohydrates, dietary fiber, vitamins, minerals, and protein.8,9 To ripen properly on the palm, dates require hot and dry weather conditions.10 Due to their nutritional value and sweet taste, dates are widely consumed as a snack and are used in various culinary preparations around the world.
Date palm fruit undergoes several stages of development before it reaches its ripening stage.11 These stages involve intricate biochemical reactions that result in the accumulation of various metabolites, including sugars, organic acids, and Phenolic compounds. These metabolites are responsible for the fruit’s distinctive flavor, aroma, and nutritional properties.12
The earliest stage of the development of date fruit is referred to as the Habauk stage, during which the fruit is small, hard, and green, and has a bitter taste. Subsequently, the fruit advances to the Kimri stage, where it becomes larger, softer, and begins to turn yellow. The Khalal stage follows, during which the fruit continues to ripen, becoming even larger, juicier, and sweeter. At the Rutab stage, the fruit reaches its peak for consumption, having attained its maximum size and being fully ripe, soft, and sweet. Finally, the fruit reaches the Tamer stage, where it has dried out and is no longer suitable for fresh consumption.11
Throughout the development of fruits, several biochemical changes occur that result in the accumulation of sugars, organic acids, and Phenolic compounds. Among the primary sugars found in dates are glucose, fructose, and sucrose, with glucose and fructose being the most abundant in ripe dates. Unripe dates derive their sour taste from organic acids such as citric, malic, and succinic acid 13, although their levels decrease as the fruit ripens. Dates also contain Phenolic compounds, including flavonoids and tannins, which possess antioxidant properties and offer potential health benefits.14
The factors influencing the chemical transformations that occur during the formation stages of dates include genetics, environmental conditions, and agricultural practices.3 This study aims to estimate and statistically test the linearity of the dynamics of nutritional compounds, acidity levels, water content, dry matter, secondary metabolites, and antioxidant activity capacity across the fruit development and maturity stages.
Materials and methods
Plant Material
The plant material used is the fruit of the date palm of the Deglet Nour and Degla Beida cultivars . Deglet Nour is a cultivar of date fruit that is known for its sweet and succulent taste, earning it the nickname “the queen of all dates”.15 it is a semi-dry type of date with slightly flaky skin and it is one of the most commercially important and widely planted date palm, originating in Algeria in the 1600s.16,17
Degla Beida is another cultivar of date palm tree that is grown in North Africa and the Middle East.18 It is a cultivar of date fruit that is dry and the least extensively planted. It is also one of the cultivars that grow in Algeria.
Collection and preparation of date samples
In this study, we selected two cultivars of local dates, Deglet Nour and Degla Beida. We chose 10 palms of each cultivar that were as homogeneous as possible in terms of age, height, and vegetative growth. We harvested the fruits using modern and sound collection methods, such as scissors and paper bags. At each stage, we collected 60 fruits, 6 from each of the 10 palms of each cultivar and divided them into three groups.”
After the sampling process, each time we cleaned the fruits with wet wipes. Then, we removed the cones and cores separately and cut the fleshy part of the fruits into very thin slices, we spread the slices on a piece of gauze with continuous stirring and dried them in a dark room. Next, we ground the dried slices automatically using an electric device and sifted the resulting powder to homogenize the granules of the dry matter of the fruits. Finally, we placed the powder in sealed glass containers until the necessary analyses were carried out.
Estimate studied chemical properties
Acidity level (PH)
The acidity level is estimated using the Marx method (1999).19
Water content (WC) and dry matter (DM) Estimate
Dry material ratio and Water content was estimated using a drying method 20, in which the fresh plant material was dried in an incubator at 60 °C. After every 24 hours, the plant material is weighed until the weight is stable. These contents calculated using the following equations:
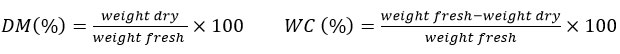
The nutritional compounds
The nutritional compounds in the fruits were estimated by extracting (protein, fats and carbohydrates) using a method of Shibko21 , from the date fruit powder, and then the carbohydrates were determined according to the method 22, while proteins were estimated according to the method 23, Whereas, fats were estimated according to the modification of the Goldsworthy meathod.24
The secondary metabolites
The secondary metabolites were estimated according to a specific method using date powder.25 The yield of extraction operations was calculated according to the following equation:
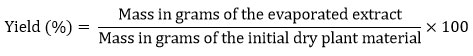
The total content of phenols was estimated according to many steps26, and the total content of flavonoids was also estimated according to steps using the Folin-Ciocalteu and the aluminum chloride colorimetric methods, respectively.27
Antioxidant activity DPPH
The DPPH radical scavenging activity of the extracts was determined using the DPPH test.28 DPPH absorbs at 517 nanometers in its radical form, but its absorption decreases upon reduction by antioxidants or radical species. To conduct the test, a 0.1 mmol L-1 solution of DPPH in methanol was prepared, and 4 mL of this solution was mixed with 1 mL of the samples dissolved in methanol at different concentrations. After 30 minutes, the absorbance was measured at 517 nanometers. A decrease in absorbance in the reaction mixture indicates higher activity in scavenging free radicals.
The antioxidant capacity to scavenge DPPH radicals was calculated using the following equation:

The sample concentration providing 50% free radical scavenging activity (IC50) was determined from the graph of DPPH scavenging effect percentage against sample concentration. Vitamin C was used as an antioxidant standard for activity comparison.
Statistical models and methods
To track the dynamics of chemical properties during growth stages, we utilized regression analysis to estimate the dynamic parameters.29 To handle potential non-normal distributions resulting from small sample sizes across study groups, we employed the generalized linear model with the Newton-Raphson method.30 This approach allowed us to estimate and test the regression function parameters effectively. We utilized the Akaike Information Criterion to determine the most suitable regression model and its corresponding independent variables for each chemical property. Our objective was to select the model from several alternative models with the lowest criterion value 31, ensuring the best fit. For each chemical property’s regression function there were alternative models, in each one we considered several potential independent variables: stage, cultivar, and their interaction (stage*cultivar) expressed as a polynomial function with varying degrees.32 And test three alternative forms: linear, quadratic, and second-order polynomial as possible functions.29,33 By selecting the best statistical regression models for each chemical property, we evaluated the direction and nature of the dynamics across growth stages. We achieved this by applying mathematical techniques, specifically the partial derivatives of each function with respect to the stage variable.34 This allowed us to study how the dynamic parameters changed throughout the various growth stages, addition determines the possible local limits (minima or maxima) of each chemical property and estimate the time of each limit.35 To track the dynamic velocity of variations in each chemical property during the growth stages and analyze their simultaneous dynamics, we calculated a standardized metric known as the relative change of chemical properties within each stage. This metric represents the change in each property in terms of standard deviation of changes of this property along all stages.36 The formula used to calculate this metric is as stated below:
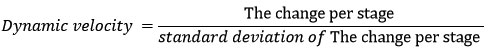
Subsequently, we illustrate them in a bar chart to enhance the comparison of dynamic velocity within the interaction of stages, chemical properties, and date cultivars.
Results
Models specification and regression results
After estimating all possible models (linear, quadratic, and second-order polynomial) by considering potential variables (stage and cultivar and their interactions), and based on the minimum value of Akaike information criterion shown in the table (1).
Table 1: Regressions selected criterion (Akaike Information Criterion)
|
Dependent variables function form |
Antioxidant activity |
yield of extraction | Total phenol | Fat
|
dry matter | Water (%) | Carbo hydrate |
Protein | PH | Total flavonoid |
| Linear | 9.0 91 |
4.975 | 11.011 | 2.484 | 7.395 | 7.806 | 9.6 61 |
5.4 53 |
0.5 81 |
5.2 |
|
Quadratic |
9.086* | 4.998 | 11.508 | 2.452 | 7.100* | 7.175 | 10.8 73 |
5.4 29 |
0.5 76 |
5.4 37 |
| Second-degree polynomial | 9.226 | 4.806* | 10.104* | 2.286* | 7.161 | 6.710* | 9.4 06* |
4.9 52* |
0.4 16* |
4.3 |
|
Coefficient Determination (R2) |
0.6503 | 0.6430 | 0.8734 | 0.9428 | 0.8609 | 0.9328 | 0.9 817 |
0.78 86 |
0.62 59 |
0.93 |
According to star marked values in the table ( 1), the appropriate independent variables and their optimal forms for each regression are as follows:
The best functional forms for Carbohydrate, Protein, Yield of extraction, and Total phenol were determined to be second-order polynomials. These functions incorporated all the independent variables, including (stage, stage2, cultivar, cultivar * stage, and cultivar * stage2). As in table (1) below, all of these independent variables had a significant effect on the chemical properties mentioned above at a significance level of 0.05, except for the variable “stage” in function of total phenol was no significant.
For water content, pH, fat content and total flavonoid content, the best form was second-order polynomial, but including different independent variables which were (stage, stage2, cultivar and cultivar * stage). Except water content function which dropped the (cultivar) variable, and pH function which dropped the (cultivar * stage) variable. All of these independent variables had a significant effect on the chemical properties mentioned above at a significance level of 0.05, except for the variable ” cultivar * stage ” in function of total flavonoid was no significant as shown in the table (1). The best functional forms for dry matter and Antioxidant activity were found to be quadratic. The dry matter function consisted of a single independent variable (stage2), while the Antioxidant activity function included multiple independent variables (stage2, cultivar, cultivar * stage2). Table (2) showed that the all of the independent variables had a significant effect on the mentioned chemical properties at a significance level of 0.05, except for the variable “cultivar * stage2” in the Antioxidant activity function, which had a significance level of 0.10.
Discussion
Dynamic analysis of selected models
pH and water content
The coefficient of determination the change of the amount of water in the fruit through the stages of growth were 93.28% table(3), The polynomial function of the second degree with a local maxima limit was the best function for the regression of the amount of water in the fruit on growth stages. before first stage, the amount of water in the date fruit was different in the two cultivar and was (57.432) ( 51.082), in Deglet Nour and Degla Baida respectively. but the tow cultivar was had the same rate of water dynamic through stages , by this rate the water firstly increased until reach its maximum in the end of second stage (56 days), which was 73.563 for Deglet Nour and 67.213 for DEGLAT BAIDA. After the second stage the water beginning decreasing at the same rate until the fifth stage where was 18.532 for Deglet Nour and 12.182 for Degla Baida. As previously mentioned, there is variation in moisture content between the two cultivars at different stages of maturity.37,38 Moisture is a crucial component that significantly impacts fruit quality and preservation. It is utilized throughout all stages of ripeness (Cambrian, Khallal, Rutab, and Tamr) in various forms, such as falling dates during natural thinning during the Rutab stage (half-ripe) and ripe dates.
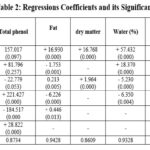 |
Table 2: Regressions Coefficients and its Significances |
These findings are consistent with previous research 39–41, that concluded soft dates have less than 30% moisture content, while dry dates have less than 10% at the date stage. The difference in moisture content between the two cultivars can be attributed to their unique genotypes.42 Confirmed the significant influence of cultivar on humidity percentage, classifying Deglet Nour as a semi-moist fruit and Degla Baida as a dry, hard fruit. The disparity can also be linked to the initial moisture percentage, fruit surface properties, exposure to climatic factors (sunlight and heat), and dry matter content.
Table 3: Dynamic local limits of chemical properties functions
|
Target measurement |
Cultivar comparisons | Water (%) |
PH |
|
Initial Stage |
DEGLAT NOUR | + 57.432 | + 5.461 |
|
DEGLA BAIDA |
51.082 |
+ 5.461 |
|
| The Difference | – 6.350 |
0 |
|
|
Rate of change over stages |
DEGLAT NOUR | + 18.370 – 10.46*S | + 0.646 – 0.172*S |
|
DEGLA BAIDA |
+ 18.370 – 10.46*S |
+ 0.562 – 0.172*S |
|
| The Difference | 0 |
-0.084 |
|
|
Local limit |
DEGLAT NOUR | 73.563 | 6.673 |
|
DEGLA BAIDA |
67.213 |
6.379 |
|
| The Difference | – 6.35 |
-0.294 |
|
|
Period of local limit |
DEGLAT NOUR | 1.76 | 3.75 |
|
DEGLA BAIDA |
1.76 |
3.26 |
|
| The Difference | 0 |
– 0.49 |
|
|
Fifth Stage |
DEGLAT NOUR | 18.532 | 6.541 |
|
DEGLA BAIDA |
12.182 |
6.121 |
|
| The Difference | – 6.35 |
– 0.42 |
The second-degree polynomial function was the best form to fit the regression of pH in date fruit on its growth stages with local maxima limit in both Deglet Nour and Degla Baida table (3). the coefficient of determination showed that 62.6% of the total pH variation in the date fruit is explained by (stage, stage2 and cultivar * stage). the difference in its growth stage. In the beginning of the first stage the pH was 5.461 in the two cultivars . but through the growth stages pH was increasing at different decline rates, in Deglet Nour the rate was while it was for Degla Baida. pH rates maximum values 6.673 and 6.379 during fourth stage(128 and 112) days, in Deglet Nour and Degla Baida respectively. In the fifth stage pH declined to 6.541 and 6.121. The change in acidity of the date fruit extract during different stages can be attributed to variations in water content, solutes, proteins, sugars, chemical elements, and secondary metabolites . These changes contribute to the fruit’s ripening process, color development, and flavor enhancement. These findings align with the results reported by in their studies on dates .43–46 Minor differences can also be attributed to genetic diversity, agricultural services and medium conditions, although these results are in contrast with the results of.
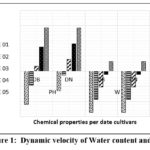 |
Figure 1: Dynamic velocity of Water content and pH |
The nutritional compounds content
Dry matter
The interpretation of the independent variable (stage2 ) in the regression accounted for 86.09% of the dry matter variance table(4). The dry matter corresponds to a second-order regression function, while neglecting any differences between the two cultivars. In the beginning of the first stage the dry matter was 16.768 in two cultivars. Through the stages of synthesis of the date fruit, the change of dry matter was positively with the progression of the stages and at an increasing rate . This means that the highest value of dry matter in the two cultivars was in the final ripening stage of dates, as it reached 65.868 in Deglet Nour while it was 68.868 in Degla Baida.
The observed difference in dry matter content between the two cultivars is attributed to the difference in the nature of the dry and semi-dry fruits. The increase in dry matter content during the ripening stages in both cultivars is due to a decrease in fruit moisture content as they progress in ripening which was confirmed by others.47 Additionally, there is an increase in the migration of nutrients and organic matter into storage tissues, particularly the fruits.48–50
Carbohydrate
The data of the change of carbohydrates through the stages of fruit synthesis fit the shape of the polynomial regression function of the second degree with a local maxima limit, with the inclusion of the difference between the two cultivars table(4). The simultaneous changes of (stage, stage2, cultivar, cultivar * stage, and cultivar * stage2) explained 98.17% of the total variation of Carbohydrates. Before the first stage, the carbohydrates in the two cultivars were different, where they were zero and 90.824 in Deglet Nour and Degla Baida respectively. Addition to that the dynamic of Carbohydrates through its growth stages were different in the two cultivars , for DEGLAT NOUR it increased at decreasing rate and at rate for Degla Baida, the two local limit was estimated out of growth stages. In the fifth stage the level of Carbohydrate was different where was 335.111 in Deglet Nour and 793.564 in Degla Baida.
The high carbohydrate content in the samples can be attributed to the fact that dates are a source of sugars, as demonstrated in numerous previous studies, such as the biochemical study of 54 cultivars of palm trees cultivated in Algeria.47,51,52 The sugar content in these studies ranged from 59.18% to 92.4%.
The results of monitoring carbohydrate content during the stages of date fruit development showed a gradual increase towards the maturity stage. This increase varied from one stage to another in the fruit of the two cultivars , as shown in Table 1. The sugar content in both Deglet Baida and Degla Nour fruits was higher than the cultivars studied by Bacha and others.53 These latter studies demonstrated that there is a difference in content and conversion rates during different stages among cultivars , which is consistent with the results of this study. This difference may be attributed to the sensitivity of regions with potential soil and air dryness, such as sandy soils with large pores that lead to water deficiency and rapid depletion.54
Studies have indicated that the onset of fruit softening is associated with fruit growth cessation and carbohydrate accumulation. This may also be attributed to the osmotic potential resulting from the accumulation of glucose 55–57, as well as growth-promoting hormones such as cytokinins 58, Cultivars also vary according to the genetic pattern at the moment and length of the cell division stage. The chemical content may differ among cultivars depending on the genetic pattern at the moment and length stage the cell division and expansion, which leads to variations in material accumulation in the fruits. This can also be explained by the elevated temperature, which increases daily consumption, explaining the slow growth and increased material content in the fruit during the initial stages.59
The significant increase in sugars in the later stages can be explained by the deterioration of complex sugars and the increasing enzyme activity during the final stages for fruit softening and ripening, where the plant’s nitrogen requirement decreases, resulting in increased sugars and carbon in the fruit . The difference between cultivars may also be attributed to variations in the efficiency of nutrient utilization, as confirmed in apple cultivars 60, and an increase in the metabolic enzyme activities of sugar, as well as a decrease in respiration rate that reduces the consumption of produced materials and consequently increases nutrient and sugar transfer to the fruit. This is accompanied by various changes in primary and secondary metabolites. Furthermore, in the late ripening stages, fruit growth ceases, starch and complex sugars degrade into soluble simple sugars, leading to gradual loss of firmness and changes in pigments.33 Studies have demonstrated the different responses of cultivars to factors influencing the carbohydrate source, including fruit load, leaf ratio, and pollination.61–64
The slow increase in carbohydrate content during the initial stages (H and K) can be explained by the utilization of the products of photosynthetic synthesis to release the energy needed for cell division, cell expansion, and tissue formation. The continued production and migration of organic matter to the fruits for storage, coupled with the low moisture content, justify the increase in sugar content during the ripening stages. This coincides with the activity of enzymes such as invertase, which converts sucrose into glucose and fructose, known for its role in cell wall softening of the fruits and the increase in sugar content, which is an indication of fruit ripening.65
Table 4: Dynamic local limits of chemical properties functions
|
Target measurement |
Cultivar comparisons | dry matter | Carbohydrate | Protein | Fat |
|
Initial Stage |
DEGLAT NOUR | + 16.768 | – 122.264 | + 24.083 | + 16.930 |
| DEGLA BAIDA | + 16.768 | 90.824 | 12.584 |
10.704 |
|
|
Rate of change over stages |
DEGLAT NOUR |
+ 3.928*S | + 318.460 – 50.788*S | – 21.523 + 7.288*S |
– 1.733 + 0.426*S |
| DEGLA BAIDA | + 3.928*S | + 204.673 – 25.65*S | – 6.045 + 2.078*S |
– 1.307 + 0.426*S |
|
|
The Difference |
0 | – 113.967 + 25.138*S | + 15.478 – 5.21*S |
+ 0.426 |
|
| Local limit |
DEGLAT NOUR |
– | 876.05 | 0 |
13.29 |
| DEGLA BAIDA | – | 907.413 | 3.8 |
8.700 |
|
|
The Difference |
– | 31.363 | 3.8 | – 8.624 | |
| Period of local limit | DEGLAT NOUR | – | 6.27 | 2.05 |
4.06 |
|
DEGLA BAIDA |
– | 7.98 | 2.90 | 3.07 | |
| Fifth Stage |
DEGLAT NOUR |
65.868 | 835.111 | 7.575 |
13.49 |
| DEGLA BAIDA | 65.868 | 793.564 | 8.358 |
9.5 |
Protein
The second-degree polynomial function was the best form to fit the regression of protein in date fruit on its growth stages with local minima limit in the two cultivars table(4). the coefficient of determination showed that 78.86% of the total protein variation in the date fruit is explained by (stage, stage2, cultivar, cultivar * stage, and cultivar * stage2). Before the first stage, the protein in both cultivars was different, as it was 24.083 in Deglet Nour, as it was 12.584 in Degla Baida. Stage by stage, the protein in Deglet Nour was decreasing at decline rate and reached the minimum level zero, daring second stages, in Degla Baida it was different where the protein was decreasing at decline rate until reached its minimum level (3.8) at the end of third stage (2.90). In last stage of growth, the protein reached 7.575 and 8.358 in Deglet Nour and Degla Baida respectively.
The variation in protein content in the fruits of the two date palm cultivars during fruit development stages is attributed to differences in climatic factors from one stage to another66, especially temperature and humidity. As the temperature increases, the plant’s consumption of organic matter also increases, leading to a reduction in the accumulation of nutritional compounds, including proteins. These conditions put the plant under stress, activating enzymes that break down primary metabolites and oxidize them through free radicals.
Additionally, the variability in the dynamics of nutritional compounds according to the cultivar is due to genetic differences and their responsiveness to environmental conditions, as well as the nature of the fruits.
These findings are consistent with the results obtained by other52 in the first three stages; however, they vary based on regional and varietal differences, as well as aligning with other findings.67,68
Fat
Regression independent variables (stage, stage2, cultivar and cultivar * stage ) explained 94.28% from total variance of fat. before dynamic table(4), Fat was 16.93 in Deglet Nour and 10.704 in Degla Baida. Dynamically the two cultivars were good fit the second degrees polynomial with local minima value, this mean that Fat in Deglet Nour and in Degla Baida were decreasing at declining rates respectively. The dynamic of fat in DEGL BAIDA were slower than Deglet Nour where reached its minimum value (8.7) in the beginning the fourth stage (105)days while Deglet Nour was late until the beginning the fifth stage (133)days to amount (13.29 ). in the final stage Fats recovered its some values for the two cultivars, in Deglet Nour was 13.49 and in Degla Baida was 9.5.
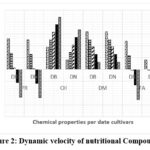 |
Figure 2: Dynamic velocity of nutritional Compounds |
The low-fat content in the studied samples can be attributed to the fact that the date palm is not a fatty plant; rather, it is an important source of sugars, with fat content ranging from 0.43% to 1.9%. These findings are consistent with several studies, such as one research69 that determined the nutritional composition of fruits from 10 date palm cultivars cultivated in Saudi Arabia, as well as the study conducted by Ahmed (1995).70 The difference in fat content between the two date palm cultivars during fruit development stages can be attributed to genetic variations and may also be influenced by differences in the source of pollination.71
Through the results of Figure 2, which illustrate the rate of changes in nutritional compounds in the fruits of the Degla Beida and Degla Nour cultivars during the ripening stages, we observe a consistent pattern across the stages for dry matter content, carbohydrate content, and protein content. The rate of change in dry matter content increases as the ripening stage advances. However, Degla Beida exhibits higher carbohydrate content compared to Degla Nour, despite both cultivars experiencing a decrease in carbohydrate content as ripening progresses. On the other hand, there is a decrease in the rate of change in protein content, followed by an increase after the third stage, reaching its peak at the fifth stage for both cultivars. Regarding fat content, the rates of change and transformations differ across the ripening stages. Degla Nour demonstrates higher rates of change in the first three stages, while Degla Beida excels during the fourth and fifth stages. These differences in the rates of change in carbohydrate and fat content can be attributed to the nature of the fruits, the length of the stage (genetic traits), and their varying impact due to changing climatic conditions from the beginning of spring to the end of autumn. Additionally, the phenological properties that distinguish each cultivar also contribute to these variations.
Secondary metabolic compounds and antioxidant activity(DPPH)
According to the results in table (5)
Yield of extraction
The regression showed that almost 64.3% of Yield variability was explained by changing of (stage, stage2, cultivar, cultivar * stage, and cultivar * stage2). before first stage the average Yield was 39.21 for Deglet Nour, and 25.322 for DEGLA BAIDA. Addition to this different, across stages the dynamic of Yield was different between the cultivars although they fit the same regression form on stage variable which was the second-degree polynomial. For Deglet Nour the Yield function had local minima value (12.512) at 120 days(stage= 3.717 ) the middle of fourth stages 140 days (4.34), before this stage point the yield decreased from stage to stage by an average , afterword the yield recovery some its level until fifth stage where reach (5.11). Whereas Degla Baida the Yield function had local higher minima value (14.568) at 140 days (stage = 4.341) almost fifth stage, before this point of time the Yield has decreased by the average according to the prior stage of change, but after that the Yield has increased until amount to (28.68) in the fifth stage.
Total phenol
The (stage, stage2, cultivar, cultivar * stage, and cultivar * stage2) explained 87.34 % of the Total phenol variability, before entering to the first stage Total phenol of Degla Baida were 378.444, greater than of Deglet Nour by 221.427. during growth stages in Deglet Nour the total phenol was increasing during first stage at a declining rate reach its maximum 230.46 at the end of the second stage 58 days, after this point of time the total phenol was decreasing at the same rate and rich to zero before the end of the fourth stage. In the Degla Baida the total phenol was firstly decreasing at diminishing rate and rich its minimum value 209.421 at the end of the second 60 days, after that the total phenol recovery its values at the same rate until reach the level 15.915 at the end of the fifth stage.
Total flavonoid
The second-degree polynomial function was the best form to fit the regression of Total flavonoid in date fruit on its growth stages with local minima limit in both Deglet Nour and Degla Baida. the coefficient of determination showed that 93.40% of the total flavonoid variation in the date fruit is explained by (stage, stage2, cultivar and cultivar * stage).
In the beginning of first stage the Total flavonoid was 9.275 in Deglet Nour and 17.761 in Degla Baida. also, through growth stages Total flavonoid in Deglet Nour were decreasing at declined rate until reached its minimum zero in beginning of second stage, while in Degla Baida were decreasing at declined rate were it reached its minimum 1.92 in the beginning of the fourth stage 112 days. after this limit total flavonoid recovered its some values intel fifth stage where it was 1.225 and 5.786 in Deglet Nour and Degla Baida respectively.
Antioxidant activity
According to the estimated regression, 65.03% of the total change in antioxidant activity can be explained by the variables stage2, cultivar * stage2, and date cultivar. In the initial first stage, there was a significant difference in the average antioxidant activity between the two cultivars at a 5% level. Deglet Nour exhibited an average of 88.076, while Degla Baida had an average of 52.343. Furthermore, the marginal effect during successive stages of growth varied between the two cultivars at a 10% level. As the fruit progressed from one stage to the next, the antioxidant activity decreased at an increasing rate, with a coefficient of (-8.522 × stage) for Deglet Nour and (-5.082 × stage) for Degla Baida, indicating that the antioxidant activity reached zero in the fifth stage of growth for both cultivars.
Table 5: Dynamic local limits of chemical properties functions
|
Target measurement |
Cultivar comparisons | yield of extraction | Total phenol | Total flavonoid | Antioxidant activity |
|
Initial Stage |
DEGLAT NOUR |
+ 39.210 | 157.017 | + 9.275 | 88.076 |
| DEGLA BAIDA | 25.322 | 378.444 | 17.761 |
52.343 |
|
|
Rate of change over stages |
DEGLAT NOUR |
– 14.720 + 3.96*S | + 81.796 – 45.558*S | – 8.435 + 2.73*S | -4.261*S |
| DEGLA BAIDA | – 4.958 + 1.142*S | – 102.721 + 12.086*S | – 9.22 + 2.73*S |
– 2.541*S |
|
|
The Difference |
+ 9.762 – 2.818*S | – 184.517 + 55.644*S | -0.785 | + 1.720*S | |
| Local limit | DEGLAT NOUR | 12.512 | 230.446 | 0 |
– |
|
DEGLA BAIDA |
14.568 | 209.421 | 1.92 | – | |
| The Difference | 2.056 | – 21.025 | 1.92 |
– |
|
|
Period of local limit |
DEGLAT NOUR |
3.717 | 1.80 | 2.7 | – |
| DEGLA BAIDA | 4.341 | 1.846 | 3.38 |
– |
|
|
Fifth Stage |
DEGLAT NOUR |
15.11 | – 3.748 | 1.225 | -18.449 |
| DEGLA BAIDA | 28.68 | 15.915 | 5.786 |
-11.182 |
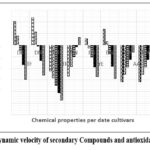 |
Figure 3 Dynamic velocity of secondary Compounds and antioxidant activity |
According to figure 3, which presents the rate of change values for extraction yield, total phenol and flavonoid content, and antioxidant activity during the ripening stages of Degla Baida and Degla Nour dates, several observations can be made.
For extraction yield, there is a decrease in the rate of change across the stages, reaching the lowest value in the fourth stage, followed by an increase in the fifth stage for both cultivars. However, Degla Baida has higher values in the first four stages, except for the fifth stage where Degla Nour excels.
Regarding flavonoid content, there is a decrease in the rate of change until the third stage, followed by an increase in the fourth and fifth stages for both cultivars, aligning with the continuous decrease in content.
The rate of change in antioxidant activity shows an increasing trend across the stages, with similarity between the two cultivars.
In terms of total phenol content, the rate of change differs between the cultivars. It decreases in Degla Baida as the fruit matures, while in Degla Nour, it decreases between the first and second stage and then increases as the fruit progresses in maturity.
These differences in the rates of change and their similarities can be attributed to the genetic nature of the cultivars within the same plant family and species. Additionally, intermediate factors between the stages, responding to the genotype and short phenological properties, play a role. The property of dry, hard, and fibrous fruits in Degla Baida, which is known for its disease resistance, may also contribute to these variations, limiting the activity of chemical transformation factors, especially enzymes.
The similarity in the rate of change of certain chemical properties between the two cultivars can be attributed to their shared family and species. However, the genetic differences between the cultivars and the source of pollen contribute to the variation in the rate of change of other chemical properties 72,73. The genetic nature of the plants plays a significant role in the vital processes that occur within them.
The nature of the dry, fibrous, and more solid fruits in Degla Baida affects the extent of chemical changes, particularly in terms of humidity percentage. The decrease in humidity has an impact on the chemical composition, especially hydrophilic compounds that are closely related to water liberation and the activity of enzymes like phenol oxidase and peroxidase.43,74,75
Additionally, both cultivars can respond to climatic changes spanning three seasons from spring to autumn, soil variations, and agricultural practices that either facilitate or hinder vital processes.76,77
At low temperatures towards the end of the autumn season, phenolic compounds can precipitate and interact with the enzyme phenol peroxidase, causing its oxidation and resulting in a decrease in the content of phenolic compounds.78
Differences can also be attributed to the phenological properties that distinguish each cultivar, including variations in the length of stages during which the date fruit undergoes ripening. For example, Degla Baida ripens in October while Degla Nour reaches maturity in December. These phenological differences interact with climatic changes, affecting the vitality of various biological interactions and life cycles of fruit pathogens.79–81
Degla Baida is known for its resistance to diseases and pests that commonly affect date fruits. This resistance limits the activity of chemical transformation factors, particularly enzymes. Consequently, the rate of changes in secondary metabolites and antioxidant activity is more pronounced in Degla Baida.
Conclusion
This research comprehensively analyzed the chemical composition and antioxidant activity of two date palm varieties, Deglet Nour and Degla Beida, across five developmental stages: Habobuk, Kimri, Khalal, Busr, and Tamr. The findings revealed distinct dynamic patterns of change in nutritional and biochemical properties influenced by growth stages, genetic factors, and environmental conditions. Key conclusions include:
Sugars increased significantly during the final two stages, contributing to the characteristic sweetness of mature dates. Polyphenol content peaked in the early stages, correlating with high antioxidant activity, which declined as the fruit matured. Moisture content decreased progressively, while dry matter increased, consistent with the fruits transitioning to drier forms. Variations in these components were less pronounced but followed cultivar-specific trends. Deglet Nour exhibited higher phenol and antioxidant levels early in development, whereas Degla Beida demonstrated resilience and distinct patterns linked to its fibrous, dry nature.
These insights underscore the significance of understanding fruit biochemical dynamics for optimizing harvest timing, storage, and processing techniques and enhancing the nutritional value of date palm products.
To build on these findings, future studies could focus on Studying the mechanisms of chemical changes during the developmental stages of date palm fruit and their consistency across different varieties, as well as examining the impact of external factors and agricultural practices on the dynamics of chemical changes.
Acknowledgement
The authors sincerely thank the farmers of the El Oued region for their invaluable contribution in permitting the monitoring of date palm production data, which greatly facilitated the progress of this study. We also extend our gratitude to the laboratories of El Oued University for providing the necessary resources to conduct laboratory analyses of the samples during this research.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The author(s) do not have any conflict of interest.
Data Availability Statement
The datasets generated and analyzed during the current study are included in this published article.
Ethics Statement
This study did not involve any data or biological material from human participants.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to Reproduce Material from Other Sources
This manuscript does not include material reproduced from other sources.
Clinical Trial Registration
This study is not associated with any clinical trial and does not require registration.
Author Contributions
- Djilani Ghemam Amara: Conceived and designed the study, contributed to data sampling and laboratory analysis.
- Noura Gheraissa: Conceived and designed the study, contributed to data sampling and laboratory analysis.
- Samir Aydi: Conceived and designed the study, contributed to data sampling and laboratory analysis.
- Ourida Benouamane: Conceived and designed the study, contributed to data sampling and laboratory analysis.
- Souraya Benaichi: Conceived and designed the study, contributed to data sampling and laboratory analysis.
- Mohammed Messaoudi: Conducted data analysis and statistical modeling.
- Ahmed Elkhalifa Chemsa: Contributed to writing and revising the manuscript.
- Zeid Alia, Hacene Laouedj: Contributed to writing and revising the manuscript.
- Nezar Cherrada: Contributed to writing and revising the manuscript.
References
- Alam MA, Syazwanie NF, Mahmod NH. Evaluation of antioxidant compounds, antioxidant activities and capsaicinoid compounds of Chili (Capsicum sp.) germplasms available in Malaysia. J Appl Res Med Aromat Plants. 2018;9:46-54.
CrossRef - Ionică ME, Nour V, Trandafir I. Bioactive compounds and antioxidant activity of hot pepper fruits at different stages of growth and ripening. J Appl Bot Food Qual. 2017;90:232-237.
CrossRef - Bae H, Jayaprakasha GK, Crosby K. Ascorbic acid, capsaicinoid, and flavonoid aglycone concentrations as a function of fruit maturity stage in greenhouse-grown peppers. Journal of Food Composition and Analysis. 2014;33(2):195-202.
CrossRef - Martínez S, Curros A, Bermúdez J, Carballo J, Franco I. The composition of Arnoia peppers (Capsicum annuum L.) at different stages of maturity. Int J Food Sci Nutr. 2007;58(2):150-161.
CrossRef - Sarker U, Oba S. Color attributes, betacyanin, and carotenoid profiles, bioactive components, and radical quenching capacity in selected Amaranthus gangeticus leafy vegetables. Sci Rep. 2021;11(1):11559.
CrossRef - Krueger RR. Date palm (Phoenix dactylifera L.) biology and utilization. In: The Date Palm Genome, Vol. 1: Phylogeny, Biodiversity and Mapping. Springer; 2021:3-28.
CrossRef - El-Juhany LI. Degradation of date palm trees and date production in Arab countries: causes and potential rehabilitation. Aust J Basic Appl Sci. 2010;4(8):3998-4010.
- Al-Shahib W, Marshall RJ. The fruit of the date palm: its possible use as the best food for the future? Int J Food Sci Nutr. 2003;54(4):247-259.
CrossRef - Ayad AA, Williams LL and Gad El-Rab DA. A review of the chemical composition, nutritional and health benefits of dates for their potential use in energy nutrition bars for athletes. Cogent Food Agric. 2020;6(1):1809309.
CrossRef - Sarraf M, Jemni M and Kahramanoğlu I. Commercial techniques for preserving date palm (Phoenix dactylifera) fruit quality and safety: A review. Saudi J Biol Sci. 2021;28(8):4408-4420.
CrossRef - Torahi A, Arzani K. Date palm (Phoenix dactylifera L.) fruit growth pattern. In: III International Symposium on Tropical and Subtropical Fruits 864. ; 2004:201-205.
CrossRef - Akhter MJ, Akhter S, Islam S, Sarker MdSH, Hasan SMK. Varietal influence on bioactive compounds and antioxidant activity in chilies during development stages. Heliyon. 2024;10(17):e37406. doi:https://doi.org/10.1016/j.heliyon.2024.e37406
CrossRef - Siddiq M, Greiby I. Overview of date fruit production, postharvest handling, processing, and nutrition. Dates: Postharvest science, processing technology and health benefits. Published online 2013:1-28.
CrossRef - Ismail Hamad IH, AbdElgawad H, and Soad Al-Jaouni SAJ. Metabolic analysis of various date palm fruit (Phoenix dactylifera L.) cultivars from Saudi Arabia to assess their nutritional quality. Published online 2015.
CrossRef - BENSANIA W, DJERIDANE A, BOURAS N. CONTROLLING THE ENZYMATIC BROWNING OF ALGERIAN DEGLET NOUR FRUIT. Research Journal of Agricultural Science. 2022;54(2).
- Guido F, Behija SE, Manel I. Chemical and aroma volatile compositions of date palm (Phoenix dactylifera L.) fruits at three maturation stages. Food Chem. 2011;127(4):1744-1754.
CrossRef - Jaradat AA. Date palm: production. Dates: postharvest science, processing technology and health benefits. Published online 2013:29-55.
CrossRef - Rahman H, Vikram P, Hammami Z, Singh RK. Recent advances in date palm genomics: A comprehensive review. Front Genet. 2022;13:959266.
CrossRef - Djilani GA, Touati D, Miloudi W, Kherraz K, Senoussi MM. BIOACTIVITY AND ANTI-OXIDANTS ACTIVITIES OF THE ETHANOLIC EXTRACT OF FRUITS AND LEAVES OF M. NIGRA AND M. ALBA FROM OUED SOUF AREA (ALGERIA). Scientific Studies & Research Series Biology/Studii si Cercetari Stiintifice Seria Biologie. 2018;27(1).
- Zaier MM, Ciudad-Mulero M, Cámara M. Revalorization of Tunisian wild Amaranthaceae halophytes: Nutritional composition variation at two different phenotypes stages. Journal of Food Composition and Analysis. 2020;89:103463.
CrossRef - Shibko S, Koivistoinen P, Tratnyek CA, Newhall AR, Friedman L. A method for sequential quantitative separation and determination of protein, RNA, DNA, lipid, and glycogen from a single rat liver homogenate or from a subcellular fraction. Anal Biochem. 1967;19(3):514-528. doi:10.1016/0003-2697(67)90242-4
CrossRef - DuBois M, Gilles KA, Hamilton JK, Rebers PA t, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28(3):350-356.
CrossRef - Classics Lowry O, Rosebrough N, Farr A, Randall R. Protein measurement with the Folin phenol reagent. J biol Chem. 1951;193(1):265-275.
CrossRef - Amara DG, Kherraz K, Nagaz K, Senoussi MM. Effect of chicken manure and organic nitrogen levels on yielding and antioxidant content of tuber potato at Algeria Sahara. Int J Agric Innov Res. 2015;4(1):17-21.
- Matkowski A, Piotrowska M. Antioxidant and free radical scavenging activities of some medicinal plants from the Lamiaceae. Fitoterapia. 2006;77(5):346-353.
CrossRef - Gheraissa N, Chemsa AE, Cherrada N. Biochemical Profile and In Vitro Therapeutic Properties of Two Euhalophytes, Halocnemum strobilaceum Pall. and Suaeda fruticosa (L.) Forske., Grown in the Sabkha Ecosystem in the Algerian Sahara. Molecules. 2023;28(8):3580.
CrossRef - Kaddour A, Amara DG, Moussaoui Y, Chemsa AE, Alia Z, Kamarchou A. Total phenolic and flavonoid contents of Mentha spicata leaves aqueous extracts in different regions of Algeria and their antioxidant, and antidiabetic activities. Tropical Journal of Pharmaceutical Research. 2022;21(9):1907-1913.
CrossRef - Chemsa AE, Derdouri S, Labbi Z. Total phenolic and total flavonoid contents of different solvent extracts of Bassia muricata (L.) Asch. and evaluation of antibacterial and antioxidant activities. J Chem Pharm Res. 2016;8(4):1317-1321.
- Huang HH, He Q. Nonlinear regression analysis. In: Tierney RJ, Rizvi F, Ercikan K, eds. International Encyclopedia of Education (Fourth Edition). Fourth Edi. Elsevier; 2023:558-567. doi:https://doi.org/10.1016/B978-0-12-818630-5.10068-5
CrossRef - Dupuy JF. Generalized Linear Models. Statistical Methods for Overdispersed Count Data. Published online January 1, 2018:33-75. doi:10.1016/B978-1-78548-266-3.50002-3
CrossRef - Portet S. A primer on model selection using the Akaike Information Criterion. Infect Dis Model. 2020;5:111-128. doi:https://doi.org/10.1016/j.idm.2019.12.010
CrossRef - Galvão RKH, de Araújo MCU, Soares SFC. Linear Regression Modeling: Variable Selection. In: ; 2020. https://api.semanticscholar.org/CorpusID:209912156
CrossRef - Bai Q, Shen Y, Huang Y. Advances in mineral nutrition transport and signal transduction in Rosaceae fruit quality and postharvest storage. Front Plant Sci. 2021;12:620018.
CrossRef - Yang XS. Chapter 13 – Partial Differentiation. In: Yang XS, ed. Engineering Mathematics with Examples and Applications. Academic Press; 2017:145-152. doi:https://doi.org/10.1016/B978-0-12-809730-4.00017-3
CrossRef - STROYAN KD. CHAPTER 20 – Maxima and Minima in Several Variables. In: STROYAN KD, ed. Calculus Using Mathematica. Academic Press; 1993:389-410. doi:https://doi.org/10.1016/B978-0-12-672971-9.50025-6
CrossRef - Umberfield EE, Bowie J, Kanter AS, Dixon BE, Tallman EF. Chapter 10 – Standardizing health care data across an enterprise. In: Dixon BE, ed. Health Information Exchange (Second Edition). Second Edi. Academic Press; 2023:237-255. doi:https://doi.org/10.1016/B978-0-323-90802-3.00024-1
CrossRef - Al-Hooti S, Sidhu JS, Qabazard H. Physicochemical characteristics of five date fruit cultivars grown in the United Arab Emirates. Plant Foods for Human Nutrition. 1997;50:101-113.
CrossRef - Iqbal M, Munir IM, Niamatullah M. Physiochemical characteristics of date palm (Phoenix dactylifera L.) cultivars at various maturity stages under environmental conditions of dera Ismail Khan. J Agric Res. 2011;49(2):249-260.
- Diboun I, Mathew S, Al-Rayyashi M. Metabolomics of dates (Phoenix dactylifera) reveals a highly dynamic ripening process accounting for major variation in fruit composition. BMC Plant Biol. 2015;15(1):1-22.
CrossRef - Farag KM. Date palm, A Wealth of Healthy Food. Encyclopedia of Food and Health (FOHE). Chapter, 00215. Published online 2016.
CrossRef - Fernández-López J, Viuda-Martos M, Sayas-Barberá E, Navarro-Rodríguez de Vera C, Pérez-Álvarez JÁ. Biological, nutritive, functional and healthy potential of date palm fruit (Phoenix dactylifera L.): Current research and future prospects. Agronomy. 2022;12(4):876.
CrossRef - Falade KO, Abbo ES. Air-drying and rehydration characteristics of date palm (Phoenix dactylifera L.) fruits. J Food Eng. 2007;79(2):724-730.
CrossRef - Chibi S, El Hadi D. The isolation and characterization of yeast strains saccharomyces cerevisiae cultivated on musts of common dates: Physiological studies of the adaptation and the resistance to ethanol. Algerian Journal of Environmental Science and Technology. 2019;5(2).
- Haider MS, Khan IA, Jaskani MJ. Pomological and biochemical profiling of date fruits (Phoenix dactylifera L.) during different fruit maturation phases. Pak J Bot. 2018;50(3):1069-1076.
- Rastegar S, Rahemi M, Baghizadeh A, Gholami M. Enzyme activity and biochemical changes of three date palm cultivars with different softening pattern during ripening. Food Chem. 2012;134(3):1279-1286.
CrossRef - Tafti AG, Fooladi MH. A study on the physico-chemical properties of Iranian Shamsaei date at different stages of maturity. World Journal of Dairy & Food Sciences. 2006;1(1):28-32.
- Ragab WS, Ramadan BR, Sorour MA, Ahmed NA. PHYSICAL AND CHEMICAL CHANGES IN FRUITS OF THREE DATES PALM (Phoenix dactylifera L.) GROWN IN SOUTH VALLEY, EGYPT. Journal of Food and Dairy Sciences. 2011;2(11):605-615.
CrossRef - Haggag L, Shahin MFM, Genaidy EAE, Fouad AA. Changes in fruit weight, dry matter, moisture content and oil percentage during fruit development stages of two olive cultivars. Middle East J. 2013;2:21-27.
- Antognozzi E, Tombesi A, Ferranti F, Frenguelli G. Influence of sink competition on peduncle histogenesis in kiwifruit. N Z J Crop Hortic Sci. 1991;19(4):433-439.
CrossRef - Nardozza S, Boldingh HL, Wohlers MW. Exogenous cytokinin application to Actinidia chinensis var. deliciosa ‘Hayward’fruit promotes fruit expansion through water uptake. Hortic Res. 2017;4.
CrossRef - Al-Farsi* MA, Lee CY. Nutritional and functional properties of dates: a review. Crit Rev Food Sci Nutr. 2008;48(10):877-887.
CrossRef - Haider MS, Khan IA, Naqvi SA. Fruit developmental stages effects on biochemical attributes in date palm. Pak J Agric Sci. 2013;50(4).
- Bacha MA, Nasr TA, Shaheen MA. Changes in Physical and chemical characteristics of the fruits of four date palm cultivars. Proc Saudi Biol Soc. 1987;10:285-295.
- Monzon JP, Jabloun M, Cock J. Influence of weather and endogenous cycles on spatiotemporal yield variation in oil palm. Agric For Meteorol. 2022;314:108789.
CrossRef - Aydi S, Sassi Aydi S, Rahmani R. Date-palm compost as soilless substrate improves plant growth, photosynthesis, yield and phytochemical quality of greenhouse melon (Cucumis melo L.). Agronomy. 2023;13(1):212.
CrossRef - Gerardeaux E, Jordan-Meille L, Constantin J, Pellerin S, Dingkuhn M. Changes in plant morphology and dry matter partitioning caused by potassium deficiency in Gossypium hirsutum (L.). Environ Exp Bot. 2010;67(3):451-459.
CrossRef - Koundouras S, Marinos V, Gkoulioti A, Kotseridis Y, van Leeuwen C. Influence of vineyard location and vine water status on fruit maturation of nonirrigated cv. Agiorgitiko (Vitis vinifera L.). Effects on wine phenolic and aroma components. J Agric Food Chem. 2006;54(14):5077-5086.
CrossRef - Shahsavar AR, Shahhosseini A. Pollen grain hormones of date palm pollinator cultivars and their relationship with hormones of different stages of ‘Piarom’date fruit growth. Sci Hortic. 2021;288:110389.
CrossRef - Curtis Jr OF. Diurnal translocation of carbohydrates into date fruits. Am J Bot. Published online 1947:388-391.
CrossRef - Wang Q, Liu C, Huang D, Dong Q, Li P, Ma F. High-efficient utilization and uptake of N contribute to higher NUE of ‘Qinguan’apple under drought and N-deficient conditions compared with “Honeycrisp’’’.”’ Tree Physiol. 2019;39(11):1880-1895.
CrossRef - Alikhani-Koupaei M, Aghdam MS, Faghih S. Physiological aspects of date palm loading and alternate bearing under regulated deficit irrigation compared to cutting back of bunch. Agric Water Manag. 2020;232:106035.
CrossRef - Alikhani-Koupaei M, Aghdam MS. Effect of number of suckers on date palm source-sink limitation tracked by physiological markers and carbon allocation responsive genes expression. Sci Hortic. 2022;304:111259.
CrossRef - Pallas B, Mialet-Serra I, Rouan L, Clément-Vidal A, Caliman JP, Dingkuhn M. Effect of source/sink ratios on yield components, growth dynamics and structural characteristics of oil palm (Elaeis guineensis) bunches. Tree Physiol. 2013;33(4):409-424.
CrossRef - Richardson AC, McAneney KJ. Influence of fruit number on fruit weight and yield of kiwifruit. Sci Hortic. 1990;42(3):233-241.
CrossRef - Al-Mssallem MQ, Al-Qarni AA, Al-Jamaan M. Dietary pattern of patients with type 2 diabetes mellitus including date consumption. J Public Health (Bangkok). Published online 2020:1-7.
CrossRef - Aurbacher J, Parker PS, Sanchez GAC. Influence of climate change on short term management of field crops–A modelling approach. Agric Syst. 2013;119:44-57.
CrossRef - Assirey EAER. Nutritional composition of ten date palm (Phoenix. Food Chem. 2009;112(2):406-411.
CrossRef - Awad MA, Al-Qurashi AD, Mohamed SA. Antioxidant capacity, antioxidant compounds and antioxidant enzyme activities in five date cultivars during development and ripening. Sci Hortic. 2011;129(4):688-693.
CrossRef - Assirey EA. Nutritional composition of ten date palm (Phoenix dactylifera L.) cultival fruits grown in Saudi Arabia by high performance liquid chromatography. Journal of Taibah University for Science. Published online 2014:1-12.
CrossRef - Ahmed IA, Ahmed AWK, Robinson RK. Chemical composition of date varieties as influenced by the stage of ripening. Food Chem. 1995;54(3):305-309.
CrossRef - ABED AKM. STUDY OF AMINO ACIDS AND FATTY ACIDS IN DATE PALM FRUIT CULTIVARS AM ALDEHIN AND BRAIME OF THREE MALE DATE PALM POLLINATERS. Journal of Basrah Researches (Sciences). 2007;33(3B).
- Ofstehage A. The gift of the middleman: an ethnography of quinoa trading networks in Los Lipez of Bolivia. Available at SSRN 1866330. Published online 2010.
CrossRef - Clegg MT, Durbin ML. Flower color variation: a model for the experimental study of evolution. Proceedings of the National Academy of Sciences. 2000;97(13):7016-7023.
CrossRef - Abdelali G, Charlotte H, Abdelmalek C, Réda YA, Ammar S, Boubekeur N. Adsorptive removal of methylene blue by low cost agricultural waste: Degla beida Dates Stones in a fixed-bed dynamic column. Res J Chem Environ. 2019;23(1):74-81.
- El-Hakim AFA, Mady E, Tahoun AMA, Ghaly MSA, Eissa MA. Seed Quality and Protein Classification of Some Quinoa Varieties. Journal of Ecological Engineering. 2022;23(1):24-33. doi:10.12911/22998993/143866
CrossRef - Besbes S, Drira L, Blecker C, Deroanne C, Attia H. Adding value to hard date (Phoenix dactylifera L.): Compositional, functional and sensory characteristics of date jam. Food Chem. 2009;112(2):406-411.
CrossRef - Kamal-Eldin A, George N, Sobti B. Dietary fiber components, microstructure, and texture of date fruits (Phoenix dactylifera, L.). Sci Rep. 2020;10(1):21767.
CrossRef - Benmeddour Z, Mehinagic E, Le Meurlay D, Louaileche H. Phenolic composition and antioxidant capacities of ten Algerian date (Phoenix dactylifera L.) cultivars: a comparative study. J Funct Foods. 2013;5(1):346-354.
CrossRef - Allam A, Djafnri K, Bergouia M, Khemissat EH, Tama M, Taleb B. Morphological and physicochemical characterisation of date palm cultivars from Ghardaïa (Southeast Algeria). Published online 2021.
CrossRef - Nguyen TBT, Ketsa S, Van Doorn WG. Relationship between browning and the activities of polyphenoloxidase and phenylalanine ammonia lyase in banana peel during low temperature storage. Postharvest Biol Technol. 2003;30(2):187-193.
CrossRef - Yahia EM. Postharvest Biology and Technology of Tropical and Subtropical Fruits: Mangosteen to White Sapote. Elsevier; 2011.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






