

Introduction
Isothiocyanates (ITCs) are the volatile phytochemicals that belong to the category of highly reactive organo-sulfur compounds and are identified as the main compounds responsible for the characteristic pungent aroma and bitter taste of the cruciferous vegetables e.g. broccoli, radish, watercress, cabbage, cauliflower and mustard.1 These are not present in the plants in their native form, but are formed when glucosinolates get hydrolyzed by myrosinase enzyme. Glucosinolates (GSLs) are the chemically stable and biologically inactive thioester compounds that are the precursors of isothiocyanates.2 Upon exposure to physical stresses such as cell disruption or mastication, these plants activate the myrosinase enzyme that hydrolyzes the glucosinolates into glucose and several unstable intermediates that undergo rearrangements and transform into secondary metabolites such as isothiocyanates, indoles, thiocyanates and nitriles.3,4,5 ITCs are known to possess several biological activities such as fungicidal, fungistatic, nematicidal, and bactericidal effects and also contribute to the anticancerous, neuroprotective, anti-inflammatory, cardio-protective and other health promoting properties in humans.6-9 Formation of isothiocyanates is largely affected by several internal as well as external factors like pH, temperature, availability of ferrous ions and presence of proteins like epithiospecifier protein.10,11 Epithiospecifier protein (ESP) is a cofactor of myrosinase enzyme that assists the formation of epithionitriles and nitriles as the breakdown product of GSLs with a terminal double bond. ESP hinders the synthesis of ITCs but it has been observed that cooking at about 60oC denatures the ESP and favors the formation of ITCs rather than epithionitriles.12,13
Nowadays, researchers are keen to study these compounds as they have been linked to prevention and cure (to some extent), some of the most cataclysmic diseases for human health. For instance, isothiocyanates are reported to react with phase I and phase II enzymes to detoxify carcinogens and excrete them out of the body, inhibit cytochrome p450, and suppress cancer cell proliferation thus preventing cancer.14,15 Apart from this, ITCs exert antioxidant, neuroprotective, and anti-inflammatory activities by regulating phase II antioxidants, reducing oxidative stress, preventing formation of reactive oxygen species (ROS), maintain redox homeostasis, enhance activity of free radical scavenger by promoting KEAP1/NRF2/ARE pathway and inhibiting nuclear translocation of NF-κB to prevent inflammation.16-19
Furthermore, various studies have confirmed the strong antimicrobial properties of ITCs as they have been reported to reduce oxygen consumption of microbes, cause oxidative stress that reduces microbial growth and alter the structure of their proteins by forming conjugates with them.20-23 Strong antibacterial and antifungal activities of some isothiocyanates such as benzyl, phenyl and allyl isothiocyanates have been reported previously.24,25 These bioactive components are considered as strong electrophiles which may easily react with nucleohilic reagents including sulphites, alcohols, amines, water and amino acids in the course of food handling as well as in physiological circumstances.26 ITCs have been reported to inhibit gram-positive and gram-negative bacteria at doses ranging from 0.02 to 200 mg/l.27,4 Being food derived compounds, ITCs are generally regarded as safe (GRAS) to be in contact with food products. These properties justify the potential of ITCs to be used as a good natural alternative for chemical antimicrobial agents in food packaging in order to improve the shelf life of different food products. The probable mode of actions of ITCs against microbial activity include disruption of various cellular mechanisms like inhibition of functioning of enzymes, alteration of structure of enzymes and increased oxidative stress.27 Moreover, it has been found to increase the shelf life of raw milk by activation of lactoperoxidases (LPO) system. Lactoperoxidases (LPO) hinders the growth of food borne micro-organisms and hence increases the shelf life of milk due to its bactericidal and bacteriostatic properties. Hydrogen peroxide assists in conversion of thiocyanate into hypothiocyanate, which is an effective antimicrobial agent due to its ability to disrupt sulfhydryl group (-SH) in bacterial enzymes. Therefore, these compounds are becoming an emerging field of interest in shelf-life extension of foods in recent decades. However, it is very challenging to extract these compounds due to their bioavailability, high volatility and heat sensitivity. Apart from that, different food processing treatments tend to influence their bioavailability resulting in alteration of their health promoting properties. Thus, in this review, an attempt has been made to explore the possible health benefits of isothiocyanates, effect on bioavailability and stability during processing, methods of extraction, taste and flavor perception and their utilization in packaging and increasing the shelf life of a variety of food products.
Types and sources of isothiocyanates
Glucosinolates, precursors of ITCs, can be found in all the parts of a plant viz. roots, seeds, leaf, or the stem. The youngest tissues possess the maximum amount of glucosinolates which decreases upon maturation due to their degradation to support the formation of other sulfur-containing compounds.28 The nature of glucosinolates in plants varies according to various factors such as climate, soil, harvesting, storage, processing, pH and plant growth conditions, for instance, sulfur fertilization favors the synthesis of aliphatic glucosinolates, whereas, nitrogen rich fertilizers favor the synthesis of indole glucosinolates.29 Individuals of the Brassicaceae family contain a variety of glucosinolates, each when hydrolyzed, shapes a distinctive isothiocyanate. The basic structure of glucosinolates comprises of a β-D-thioglucose group, a sulfonated oxime group and an amino acid based side chain.30 Depending upon the configuration of the chain, glucosinolates may either be aliphatic, x-methylated, aromatic, or heterocyclic (indole).31 Since the isothiocyanates are hydrolysis product of glucosinolates therefore they are based on the sorts of their glucosinolate precursor and are categorized into three groups i.e. aliphatic, terpenoids, and aromatic ITCs. There are some other isothiocyanates that are synthetic in nature, for example, methyl isothiocyanate, propyl isothiocyanate, furfuryl isothiocyanates, and phenyl isothiocyanates. Chemical structures of some isothiocyanates are shown in Figure 1.
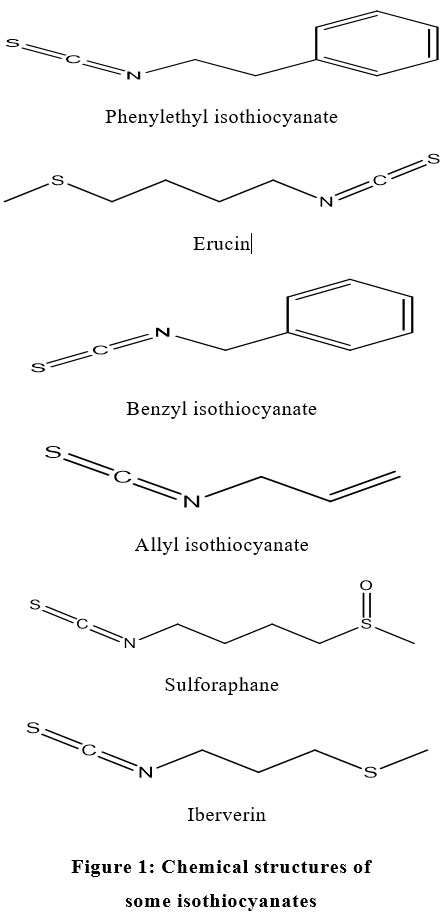 |
Figure 1: Chemical structures of some isothiocyanates. |
Chemistry of isothiocyanates
On the structural basis, isothiocyanates (ITCs) have highly reactive organo-sulfur group and are the esters of isothiocyanic acid (H-NCS) that can also be regarded as sulfur analogs of isocyanates (R-NCO).3 Their fundamental structure is, R-N=C=S, where, R can be any aliphatic or aromatic side chain while the NCS group has negative dipole due to which they readily form a wide class of nitrogen, sulphur, and oxygen heterocycles. The carbon atom of NCS group in ITCs is highly electrophilic in nature (–N=C=S), and attacks the thiols to form reversible thiocarbamates and also produces stable thiourea derivates by reacting with amines in the human body; for example, glutathione, a thiol, conjugates with ITC by catalysis of glutathione S-transferases (GSTs). This reaction acts as an important metabolic route of accumulation as well as detoxification in human system.27 Another role of isothiocyanate’s electrophilic nature is that it can covalently modify proteins which can be further utilized in laboratories for tagging proteins with other compounds such as fluorescein ITC.1 Due to their versatile chemical properties, the bioactivities of isothiocyanates include the prevention of formation of cancer cells by induction of apoptosis and production of cytokine alongwith anti-inflammatory, antimicrobial and cardio-protective properties.17
Extraction of isothiocyanates
The extraction of isothiocyanates is difficult because they are unstable and heat sensitive so require technical considerations and thorough study. Researchers have used various extraction techniques such as microwave assisted extraction, liquid- liquid extraction, supercritical fluid extraction, pulsed electric field extraction and ultrasonication for the extraction of isothiocyanates (Table 1). But these methods are under constant investigation for the improvisation in their extraction capacity without affecting the final yield and stability of ITCs. The effect of these processing technologies on the extraction of glucosinolates and isothiocyanates is discussed below:
Conventional methods
Isothiocyanates can be obtained by enzymic hydrolysis of glucosinolates in laboratories, but for doing so, it becomes necessary to extract intact glucosinolates from plant tissues. The most common processes used for extraction of glucosinolates are hot water extraction and extraction using aqueous organic solvents. Nakamura32 investigated the application of conventional distillation for the extraction of BITC from green papaya. They used reverse phase HPLC-UV at 254 nm for analysis and reported that green papaya contained about 85.6% BITC out of which, about 80% was extracted in the distilled water. But when stored at 4oC, BITC present in the distilled water was rapidly diminished over time due to the sensitivity to the temperature. Further, hydro-distillation sometimes causes burning of the material in bottom of the flask, so magnetic stirrer is preferred to prevent the material to be extracted from settling in the bottom of flask.33 Apart from this, the extraction of BITC with organic solvents such as chloroform, acetone, and ethanol has also been studied. It was reported that extraction with chloroform gave highest yield followed by acetone and then ethanol, the reason being, ethanol is hydroxylated solvent which reacts with BITC whereas acetone is miscible in water which facilitates the contact between N=C=S group of BITC and hydroxyl group of polar solvents to react with each other.34 Even though these methods are used extensively, they still possess limitations for their application, such as these require large amount of solvents, cold extraction conditions, inactivation of myrosinase as well as long extraction times.35 To overcome these hurdles, various non-conventional technologies like pulsed electric field, ultrasonication, microwave assisted extraction and high pressure processing are now being applied.
Table 1: Extraction and identification of different isothiocyanates from different natural sources.
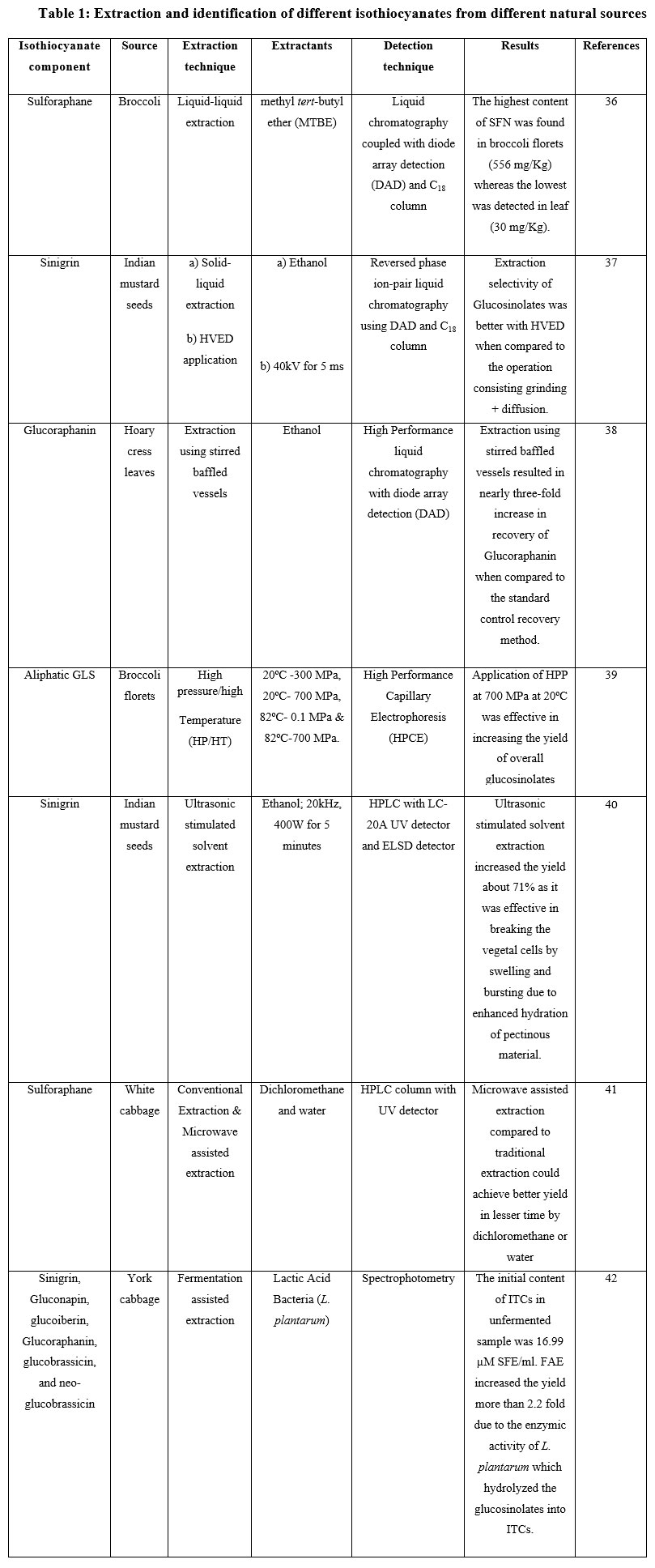 |
Table 1: Extraction and identification of different isothiocyanates from different natural sources. |
High pressure processing (HPP)
In high pressure processing, the materials are exposed to a high pressure (300-600 MPa) at ambient temperature for 1 to 15 minutes for pasteurization or sterilization usually, but now it can also be applied for the extraction of ITCs as it has proven its efficiency in enhancing the hydrolysis of glucosinolate to ITCs. For instance, in an experiment it was observed that application of high pressure processing at 500 MPa on brussels sprouts increased the concentration of sulforaphane up to 317% compared to control, because HPP caused the cells to disrupt and release myrosinase which hydrolyzed glucoraphanin to sulforaphane.43 Likewise, Westphal44 studied the effect of HPP on bioactivation of glucosinolates to isothiocyanates in broccoli sprouts. They applied 100-600 MPa pressure on Broccoli sprouts for 3 minutes at 30oC and it was observed that use of pressure at about 600 MPa increased the degree of conversion of GSLs into their respective ITCs up to 85% which was 4-5 times higher than samples treated at low pressures. This can be attributed to the inactivation of epithiospecifier protein as well as decomparmentalization of myrosinase enzyme. Overall, this method of extraction possesses no serious drawbacks but the only limitation is that high pressure raises the temperature of the material which can affect the yield of the obtained isothiocyanates or glucosinolates.
Ultrasound assisted extraction
Ultrasound is an innovative processing technology that is based on the phenomenon of acoustic cavitation i.e., rapid expansion and contraction of bubbles of gas/vapours. This generates intense local heating and high pressures that cause cell wall disruption, solvent penetration into material and also release the components of cell in food matrices. Wang40 used ultrasonication as the pre-treatment method for the solvent based extraction of sinigrin from Indian mustard seed. It was observed that without ultrasound, conventional extraction method resulted in lower yield of sinigrin (2.25 ±0.05%) whereas, ultrasound assisted extraction technique resulted in higher yield (3.84±0.02%), i.e. 71% increase in productivity. Ultrasonication resulted in elimination of external pectous substance which led to the enhanced productivity of sinigrin. Moreover, it was also reported that as the content of pectous material decreased, the content of sinigrin increased suggesting the correlation of sinigrin productivity to the amount of the pectous substance in seeds. Ultrasound treatment is also used as a pre-treatment step prior to microwave assisted extraction.
Microwave assisted extraction (MAE)
MAE is a method that involves traditional extraction technique in combination with microwave heating to increase the degree of extraction by promoting uniform transfer of heat and mass in the material with the reduced extraction time. Tanongkankit41 applied MAE for the extraction of sulforaphane from cabbage and observed that MAE, when combined with conventional extraction, resulted in a higher extraction yield in dichloromethane solvent than water because sulforaphane possesses weak polarity due to which it can easily be extracted by non polar organic solvents such as dichloromethane. Moreover, application of microwaves activates myrosinase which is an important factor for the formation of isothiocyanate. At present, Microwave assisted extraction is not in much application as it requires optimization and more detailed study.
Supercritical fluid extraction (SFE)
In supercritical fluid extraction, inert fluids above their critical values are used for the extraction.45 Advantages of this method are high selectivity, rapid extraction, non toxic and inert solvent, low consumption of solvent as well as absence of corresponding residues in the extract. Li46 optimized supercritical extraction using CO2 (SCE) for the extraction of AITC from wasabi (Wasabia japonica Matsum) and observed that the highest yield obtained was 0.40% at 25 MPa and 35oC. They also inferred that the yield of AITC increased with increasing the pressure (15-25 MPa) and varying the temperature range (35-55oC). SCF has high diffusivity due to the dense gaseous nature of the supercritical fluid that promotes higher yield of extract, thus can be used for the extraction of ITCs. Due to structural variations of ITCs, their diffusivity in SCF is variable. So, to enhance the diffusion, some modifiers such as ethanol, hexane, dichloromethane and ether are used.
Pulsed electric field assisted extraction
Although, High pressure processing and supercritical fluid extraction methods are efficient for extraction but these require high cost for the set up. In addition, HPP is so powerful in rupturing the cells that it somehow leads to leaching and formation of some unwanted compounds along with the desired phytochemicals. Therefore, a milder technology like pulsed electric field (PEF) based on the principle of electroporation through the application of high intensity electric pulses (10-80 kV/cm) for very short durations of time, can be utilized for extraction of ITCs. The principle phenomenon of electroporation of PEF processing renders the cell permeable and leads to leaching of different compounds. In a study, variables of pulsed electric field treatment (electric field strength varying from 1 to 4 kV/cm and pulse duration ranging from 50 to 1000 µs at 5 Hz) were applied on broccoli stalks and flowers to extract glucosinolate. The optimum PEF conditions reported to be adequate for the extraction of GSLs were 4kV cm-1 for 525 and 1000µs as this led to a two-fold increase in final extracted yield of neoglucobrassicin, glucoiberin and glucobrassicin in broccoli flower and stalk.47 Application of PEF to a higher degree also favors the conversion of glucosinolates into ITCs as electroporation leads to permeabilization and release of myrosinase which then hydrolyzes the glucosinolate to produce ITCs.
Fermentation Assisted Extraction
Fermentation assisted extraction is one of the latest techniques that is gaining popularity for extraction. Jaiswal and Ghannam42 applied this method by using Box-Bhenken Experimental design to extract isothiocyanates from York cabbage. During the investigation, they evaluated total initial content of GSLs to be 14.06 ± 0.074 µg/mL which consisted of seven different glucosinolates out of which, majority were aliphatic in structure. They further inactivated the myrosinase enzyme by blanching for about 12 min at 95oC before fermentation and then performed the LAB fermentation with an S/L ratio of 0.25 (w/v) for 36 hours at the agitation rate of 100 rpm. This resulted in an overall increase upto 2.2 times the initial concentration of ITCs (Sulforaphane equivalent/ml).
Thus it can be stated that the recent advancements and applications of novel technologies for the process of extraction are leading to better yields while saving energy and time. However, these methods still lack the proper research due to which these methods are not yet commercially applied for the extraction purposes but further analysis, experimentations and developments in utilization of these methods can overcome such obstacles.
High voltage electric discharge (HVED)
HVED assisted extraction is an innovative and efficient method which represents the green extraction. This is a novel alternative to traditional extraction with several advantages such as low energy input, short treatment time and higher recovery of bioactive components48. The principle of action of high voltage electric discharge is electrohydraulic discharge. This is a liquid phase discharge technology in which electrical expulsions are produced directly in water inducing several physical and chemical reactions in water. In a study, the effect of HVED treatment on the recovery of isothiocyanates, proteins and phenolic compounds from rapeseeds and rapeseed press cake was evaluated. The raw materials were subjected to different energy inputs ranging from 0 to 400 KJ/Kg and different solvent to solid ratios (1:5 to 1:20, w/w). It was concluded that high voltage electrical discharge treatment was highly effective in extracting high-added value components namely isothiocyanates, proteins and phenolic compounds49.
Pressurized fluid extraction (PFE)
PEF, also called accelerated solvent extraction (ASE) is a sample preparation technique for extracting several components from a solid or semisolid matrix with organic solvents near their supercritical region to enhance the rate and efficacy of extraction procedure. Pressurized fluid extraction reduces the requirement of high solvent volume and extraction time by using elevated temperature and pressure that allows high solubility and diffusion rate of extracts along with high penetration rate of the solvent in sample. In a study, Rodrigues50 optimized the extraction of isothiocyanates and polyphenols using pressurized fluid under controlled pressure and temperature from watercress. The pre-treatment included different conditions of incubation time varying from 0 to 120 minutes, temperature (25-35℃) and pressure at 25MPa. Supercritical carbon dioxide as well as combination of carbon dioxide and ethanol were used to recover polyphenol-enriched isothiocyanate extracts (35℃ temp; 25 MPa pressure). The authors reported that supercritical carbon dioxide promoted the extraction of isothiocyanates from watercress while the carbon dioxide-ethanol mixture isolated polyphenols.
Stability of isothiocyanates (ITCs)
These phytochemicals can easily get affected by various factors like pH, temperature, heat, and other chemical compounds. Therefore, the evaluation of stability is vital for rendering their commercial applications in various fields. Luang-In and Rossiter51 were the first to investigate the stability of ITCs and nitriles in various buffers (pH 7.0) such as citrate phosphate buffer and Tris-Cl that were incubated anaerobically at 37oC for more than 24 hours in nutrient broths and other aqueous solutions with/without the presence of bacterial cells along with the stability of those ITCs without bacterial cells. They observed that all the tested ITCs including AITC, BITC, PITC, IBV, and ERN were unstable within the tested aqueous solutions and their concentration reduced from 1.0 mM to 0.2-0.4 mM within 8 hours in nutrient broths without bacterial cells. The rationale they stated was that the nutrient broth consists of yeast extract, beef extract, peptone and NaCl that exhibit affinity to bind with isothiocyanates.26,52 Thus, they concluded that ITCs are unstable in aqueous solutions because the functional group in ITCs has electrophilic character that facilitates ITCs to react with nucleophilic agents such as water, carboxylic acids and amino acids.53
Other factors that govern the stability and reactivity of ITCs include steric hindrance, electrical effect and pKa value of the molecules in buffers. Pressure also regulates the stability of myrosinase and isothiocyanates. For instance, pressure range from 100-600 MPa (10-60oC) decreases up to 88% the activity of myrosinase but application of high pressure at mild temperature (600-800 MPa) enhances the stability of ITCs. These kinetic results confirm that ITCs have higher HP/T stability than myrosinase and hence, can be an alternative to thermal processes.54
The latest and most promising method of stabilization of isothiocyanates is microencapsulation. Encapsulation is a process that encloses a substance i.e. active agent into a secondary material and produces material in smaller size range (nm, µm or mm). The inner material is called the “core” and the outer material covering the core is referred as “wall material”, “encapsulating agent”, “coating”, “membrane”, “shell”, “capsule”, “carrier material”, “external phase” or “matrix”.55 Microencapsulation can also be used for enhancing the stability of isothiocyanates as they are volatile, heat, pH and temperature sensitive, for instance, SFN encapsulated in hydroxypropyl-β-cyclodextrin (HP-β-CD) stabilizes SFN and prevents its degradation even at temperature 50oC or reduces its loss to about 11.3% in pH 8.56 The solubility of microencapsulated ITCs such as AITC and PITC in water was investigated by Neoh57 who used randomly methylated β-cyclodextrin (RM-β-CD) to encapsulate the AITC and PITC by spray drying method to form the complexed powders for their thermal analysis. AITC and PITC had different solubility coefficient and AITC demonstrated 26-folds higher solubility than PITC and the solubility of both increased as the level of RM-β-CD was increased. This indicated the solubilization influence of RM-β-CD on the slightly soluble molecules. They also noted that even though the solubility factor of AITC was greater than PITC but RM-β-CD was seemingly more efficacious in solubilization of PITC than AITC. Further, the results of their stability studies revealed that the AITC had higher affinity to the complex than the PITC as the apparent stability constants, K1:1 and K1:2 for the AITC system were higher than that of PITC because alkyl group of AITC provides lesser steric hindrance in comparison to the benzene ring of PITC.
These investigations suggest the effective utilization of microencapsulation and non thermal technologies in preventing destabilization or degradation of volatile and thermolabile isothiocyanates. Although the techniques in practice are potent but they need further innovations and research to enhance their applicability in stabilization of compounds like isothiocyanates that are volatile and difficult to extract.
Bioaccessibility and bioavailability of ITCs in human body
Bioaccessibility is the concentration of a molecule that is released in the digestive system from a food material and is available for absorption whereas bioavailability represents the amount of administrated compounds reaching to circulatory system after being absorbed and is used for physiological functions.58,59 Being phytochemicals, isothiocyanates cannot be synthesized in situ in human or animal bodies. Therefore, the bioaccessibility of isothiocyanates in human digestive system depends on the various processing conditions applied to the vegetables which have significant effects on glucosinolate concentration, myrosinase activity and epithiospecifier protein activity.60 Cooking tends to denature the myrosinase due to its sensitivity towards high temperature and heat; therefore, when the food is cooked, some amount of myrosinase gets inactivated. Eventually, during the consumption, intact glucosinolates present in the ingested food get partially absorbed in the stomach and the rest of the glucosinolates move down through the gastrointestinal tract to the small intestine where the plant myrosinase enzyme hydrolyses them and the breakdown products thus obtained get absorbed. If some amount of glucosinolates is left un-hydrolyzed, then it is transported to the colon where bacterial myrosinase hydrolyzes them into breakdown compounds which are then absorbed or excreted.61,62.
Effect of various processing methods on bioavailability of isothiocyanates
Various treatments used in preparation, processing and storage of cruciferous vegetables may affect the bioavailability of their GSLs and ITCs compounds resulting in alteration of their health-promoting activities. The solubility of glucosinolates is an important aspect to determine their bioavailability because GLs and their breakdown products are soluble in water and therefore, when the vegetables are processed i.e. cooked or boiled; the amount of glucosinolates reduces upto 18.1 to 59.2%.63,64 The rate of conversion of ITCs is higher when consumed raw rather than after cooking the vegetables. In a study, it was observed that the rate of conversion of uncooked watercress was about 17.2 to 77.7% whereas it was only 1.2 to 7.3% in cooked watercress.65 It could be due to the reason that cooking inactivates the myrosinase enzyme present in food due to which glucosinolates reach down to the colon where they are then converted to breakdown products with the help of gastrointestinal microflora. However, the rate of conversion is slower than that of enzyme present naturally in the food.66
Apart from this, cooking methods that require high temperatures can cause thermal degradation of vegetables, for example, boiling may result in glucosinolate loss up to 5-20% as the concentration of loss is controlled by several factors such as amount of water used for cooking, boiling time, structure of vegetable tissues and the method used to cut the vegetables.67 Process of steaming can be an alternative to boiling as it prevents the leaching of glucosinolates in cooking water while increasing the concentration by enhancing the rate of extractability due to heating. A study showed that steaming of broccoli increased its total glucosinolate content up to 20-30% and steaming for shorter time inactivated ESP while increasing the formation of ITCs over nitriles.68 Besides steaming, another frequently used food preparation method is stir frying in which the cut vegetables are pan fried for a few minutes by using a small quantity of preheated oil. Stir frying limits the loss of glucosinolates by leaching as it does not require water, however, it may cause some degree of thermal degradation if the frying temperature gets too high. Various studies have reported the full retention of glucosinolates after stir-frying as it inhibited the myrosinase activity rapidly without affecting glucosinolate content.60,69-71 Other processing techniques e.g. air drying, HPP and PEF, utilized by the food industry have also been reported to have an effect on the glucosinolate-myrosinase system.44,47
Likewise, non-thermal treatments can also influence the Glucosinolate-Myrosinase system. For example, fermentation is the oldest known traditional processing method in which vegetables like cabbages are fermented in closed vessels to prepare sauerkraut and kimchi. In previous investigations, fermentation was reported to reduce the glucosinolate-myrosinase system where 7 days of fermentation led to absolute degradation of glucosinolates in white cabbage and a 13-fold reduction was noted in nozawana leaves.72,73 The possible reason for this reduction may be the extended steeping duration in water that could have possibly resulted in glucosinolate breakdown by myrosinase enzyme.72
Storage conditions are also one of the factors that affect the availability of glucosinolates. For the long-term storage of perishable products, freezing is done at a temperature around or below −20oC. Usually blanching is applied before freezing the vegetables to inactive the enzymes that can cause the quality deterioration in food products stored a sub-zero temperature. Low storage temperatures largely influence the availability of glucosinolates and isothiocyanates because during freezing, ice crystals are formed in cellular spaces that may promote cell lysis and cause hydrolysis of glucosinolate by myrosinase activity during thawing. In a study, the effective stability effect of freezing and blanching on glucosinolate content was reported in which storage of blanched-frozen broccoli and cauliflower at -20oC retained myrosinase and glucosinolates upto 90 days, whereas freezing of vegetables (−85oC for 7 days) without any pretreatment like blanching caused 1/3rd destruction of glucosinolate levels due to freeze-thaw fracture of plant cells.69 Another method of storage, i.e. cold storage at 4–8 °C for 7 days was reported to reduce the GSL levels to highest extent (1/4th) in broccoli, while lowest reduction for cabbage (1/10th) among the different vegetables belonging to cruciferae family.70
Post harvest packaging conditions may also cause fluctuations in glucosinolate content. Rangkadilok70 reported that storage of broccoli in open air caused 55% decrease in glucoraphanin content within 3 days, whereas storage in plastic bags caused 56% reduction after 7 days. The higher degradation in open air storage was most probably due to the loss of cellular structure of cells that resulted in myrosinase hydrolysis of glucoraphanin whereas during the storage in plastic bags, low oxygen level accelerated anaerobic respiration that caused cell death and spoilage of broccoli within 7 days. However, application of modified atmospheric packaging (MAP) effectively retained the glucoraphanin level for more than a week when stored at 4oC whereas controlled atmospheric storage proved to be best storage condition as it retained glucoraphanin concentration up to 25 days as it reduced the rate of degradation of broccoli florets. These findings correlate well with the investigation of Rodrigues & Rosa,75 who suggested that storage of broccoli florets for 5 days at 20oC caused 82% reduction in glucoraphanin while at 4oC, only a 31% reduction marked was observed.
Taste and Flavor Perception of Isothiocyanates
Isothiocyanates are the major compounds that are responsible for the typical sharp and pungent flavors of Brassicaceae family of plants. Apart from isothiocyanates, other degradation products of glucosinolates such as nitriles, iso-nitriles and thio-sulphur compounds also contribute to the distinctive tastes and flavors of cruciferous vegetables. Other than the natural flavors, the technological processes such as cooking, steaming, frying, and drying also substantially modify the specific flavor of these vegetables. These processes activate the various enzymatic reactions and formation of components depending upon the thermal stability of epithiospecifier protein and myrosinase enzyme that play a significant role in flavor development. For instance, Isothiocyanates influence the taste receptors such as TRPA1 and TRPV1 by contributing to the pungency of cruciferous plants. D’ Auria76 reported a reduction in pungent flavor of horshradish after grating and during refrigeration which might be attributed to the hydrolysis of glucosinolates as well as cell damage. Fats and oils control the degradation of isothiocyanates, thus can be used to maintain the pungency.77 Scientifically, bitter tastes are triggered when ingested food contains molecules that bind to G protein-coupled T2R38 receptors present on the cell membranes of taste buds. This activates the second-messenger transmitter compounds within the taste cells ensuing some intracellular chemical changes and thereby, activating the taste signals.74 Glucosinolates and isothiocyanates form covalent bonds with cysteine residue which is important for the activation of bitter taste receptors.79
Health benefits of isothiocyanates
Scientific studies have shown that regular intake of isothiocyanates is correlated with health beneficial effects such as anticarcinogenic, neuroprotective, antidiabetic, cardioprotective antioxidant, and antimicrobial effects (Table 2).
Role of isothiocyanates in cancer prevention
Cancer is one of the most noxious diseases in the world in which the transformation of normal cells into atypical, or dysplastic cells takes place that later, turns into malignant cells.80 It is stimulated through genetic as well as epigenetic changes which interrupt the pathways that control the processes of cell proliferation, apoptosis, differentiation, and senescence.81 Various factors such as genetic mutations or smoking, contact with carcinogenic compounds, obesity, radiations and some viruses like hepatitis B virus, Epstein-Barr virus, and hepatitis C virus are the primary causative agents of cancer.82 Apart from these factors, inflammation which is a common cell response can be a factor of tumor progression as it is induced by transcription factor nuclear factor-kappa B (NFκB) that controls cell proliferation by invigorating transcription of cyclin encoding genes. ITCs suppress the phosphorylation of the inhibitory proteins 1κBα and 1κB Kinase to restrain the nuclear translocation of NF-κB and prevent inflammation.19 Normal cells contain certain group of enzymes such as phase I and phase II enzymes to prevent carcinogenesis and maintain proper functioning of the cell. Phase I enzymes include monooxygenases (e.g. Cytochrome p450), dehydrogenases, estrases, reductases and oxidases which perform the function of introducing the functional groups into their substrates often resulting to form pro-carcinogens as well as convert them to epoxides which are highly carcinogenic in nature or make the compounds more hydrophilic causing increased reactivity and DNA damage. Whereas, Phase II enzymes like glutathione S-transferase (GST), sulfotranseferases, NAD(P)H oxidases, quinine oxidoreductase (NQO) etc, are responsible for detoxification as well as excretion of carcinogens.83 Some of the Phase II enzymes, for example, glutathione S-transferase, metabolize the products of phase I enzymes to form inactive hydrophobic conjugates which are then excreted in the urine.84 It has been suggested that regular intake of chemo preventive agents can reduce the chances of cancer and are divided into two groups: anti-mutagenic and anti-proliferative based on their anticancer effects. Antimutagens act to prevent carcinogen uptake or can block the activation of procarcinogens by cyclooxigenases and lipooxigenases and antiproliferative agents like topoisomerae II inhibitors (thiones such as oltipraz, phenols such as genistein) prevent the proliferation of cancer cells.85 Various fruits and vegetables contain chemo-preventive phytonutrients for example genistein, resveratrol, diallyl sulfide, lycopene, curcumin, ursolic acid, catechins, isothiocyanates, silymarin and eugenol. These compounds can suppress the propagation of cancerous cells, prevent growth factor signaling pathways and induce apoptosis.86 Wattenberg was the first person to report cancer inhibiting property of isothiocyanates.87 These are probable inducers of phase II enzymes at transcription level with the help of antioxidant response element (ARE) present in upstream region of 5’ end of mRNA. This response element is activated when nuclear transcription factor erythroid 2-related factor2 (NRF2) is released from cytosole and translocated to nucleus where it dimerizes with small Maf transcription factor and attaches to these ARE (antioxidant response element) for the initiation of transcription process of phase II enzymes.88 For instance, SFN is known to be a potent activator of phase II enzymes, inducer of breaks in DNA double strands thereby promoting cellular death, and inducer of caspase mediated apoptosis in bladder and prostate cancer cells. It is also responsible for the over expression of bax & down regulation of Bcl2.89 In addition to these properties, ITCs are also capable of inhibiting cytochrome p450, the family of enzymes which is responsible for the metabolism of all the phase I carcinogens. Furthermore, ITCs are known to prevent cancer by various mechanisms such as, 1) by modifying carcinogen metabolism, 2) initiation of apoptosis and halting cell cycle, 3) inhibition of angiogenesis and metastasis, and 4) modification in status of histone acetylation due to their property of inducing genomic alterations through, a) Nrf2 mediated induction of phase II detoxification enzymes, b) activation of cell cycle growth arrest or, c) apoptosis.16,90
Isothiocyanates as antioxidants
ITCs are the phytochemicals that promote detoxification of body and hence keep diseases at bay. These can operate as indirect antioxidants by activating the Nrf2 dependent pathway that is crucial for the regulation of phase II antioxidants and detoxifying enzymes which further eliminate the carcinogenic intermediates and protect the cells from various damages that may generate oxidative stress.91 Some ITCs like SFN behave like indirect antioxidant for their proficiency in activating several antioxidant enzymes (AO) that help in maintaining redox potential and activity of free radical scavengers such as vitamins A, C and E through KEAP1/Nrf2/ARE pathway.18 Whereas, Erucin acts as direct as well as indirect antioxidant because it possesses hydroperoxide scavenging activity due to its ability to react with H2O2 and alkyl hydroperoxides to form water and an alcohol, and indirect activity by inducing enzymes like thioredoxin reductase1 (TrxR1) in human breast cancer MCF7 cells that act as cellular antioxidants.92,93
Isothiocyanates as neuroprotective agents
Many diseases that are common to old age such as Alzheimer’s, epilepsy, brain tumor, and Parkinson’s are related to dysfunctional BBB.94 Alzheimer’s disease (AD) is a progressive neurologic disease in which accumulation of amyloid β-peptides, and intraneuronal deposition of neurofibrillary tangles composed of hyper-phosphorylated tau protein in brain take place.95 Whereas, in Parkinson’s disease (PD) loss of dopaminergic neurons occurs, along with the autooxidation of dopamine that results in formation of neurotoxic compounds like ROS, hydrogen peroxides, electrophilic dopaminergic quinone which further activate the α-synuclein associated neurotoxicity.96 Isothiocyanates have neuroprotective properties for prevention and treatment of such diseases because these target the Nrf2 pathway as it regulates some genes that are associated to induce neurodegenerative conditions.97 Along with it, these have property of reducing oxidative stress as they act as ROS scavengers and restore the balance between excessive production of ROS and antioxidant deficiency. For instance, in Okadaic acid (OKA) treated rats SFN decreased the formation of pro-oxidant species like ROS, reactive nitrogen species (RNS), TNFα and NFκB, and inhibited neuronal programmed cell death in some tissues of animal’s brain while up regulating Nrf2 and HO1 enzymes in tissue samples.98 These observations highlighted the neuroprotective properties of SFN. Furthermore, it has also been identified that SFN has the ability to inhibit NFκB pathway that might promote growth of axon after injury as its inhibition reduces the inflammation as well as chronic gliotic response which leads to axonal development.99 All these studies confirm the relation between isothiocyanates and neuroprotection to prevent neurodegenerative diseases.
Isothiocyanates in prevention of Obesity
Obesity is the condition of accumulation of excess body fat and is related with metabolic disorders. It is commonly followed by other diseases such as high blood pressure, blood cholesterol, high triglycerides level and cancer. There are many ways by which cells prevent fat accumulation for example, by secreting inflammatory mediators and preventing conversion of pre-adipocytes into adipocytes which store fat, or by regulating lipolysis with the help of negative regulators such as Galectin-12 (Gal-12) which is a β-galactoside binding protein located on large lipid droplets.100 Lowering the level of Galectin-12 impairs their differentiation into adipocytes. There are many reports linking the consumption of cruciferous vegetable extracts to reducing obesity. In one such experiment, when 31 adult volunteers were administered two cans of canned mixed green vegetable and fruit beverage containing broccoli and cabbage (160ml/can) per day for 3 weeks, it led to reduction in serum cholesterol and LDP.101 Apart from this, administration of cabbage-methanol extract in alloxan induced diabetic rabbits significantly reduced blood glucose, LDL and total cholesterol and increased High Density Lipoprotein (HDL) level.102 Furthermore, AITC was reported to suppress the concentration of Galectin- 12 and reduce formation of adipocytes thus it was concluded that ITCs could be beneficial for prevention and treatment of obesity.103
Isothiocyanates as cardio-protective agents
It has already been discussed that ITCs are effective in reducing LDL and total Cholesterol level and increasing HDL level which is an important factor for proper functioning of heart therefore they may provide protection to heart against various diseases. An isothiocyanate named 4-Carboxyphenyl isothiocyanate (4-CPI) has been shown to induce vaso-relaxation effects on coronary artery along with membrane hyper polarization on vascular smooth muscles. It is due to the activity of 4-CPI as H2S donor which helps in activation of mitochondrial K+-ATP channel.104 Another isothiocyanate SFN from broccoli is linked to reduce LDL level and increase HDL level in human blood. The main mechanism of action behind cardio-protection of ITCs involves activation of AKT/Protein Kinase B and extracellular signal regulated Kinase 1 and 2 (ERK 1/2) signaling pathway that are important for cardiac cell survival and inhibition of phase I enzymes, angiogenesis and inflammation that prevent cardiac diseases.3
Isothiocyanates as anti-diabetic agents
Diabetes often scientifically referred as Diabetes mellitus, is an ailment in which the person suffers from high blood glucose levels, either because the production of insulin in body is inadequate, or because the cells are unable to react to the insulin.105 In order to study the effectiveness of isothiocyanates in preventing diabetes, Sahin106 studied the effect of AITC on insulin resistance and transcription factors in diabetic rat model in which they observed significant reduction in blood glucose, total cholesterol, triglycerides and creatinine levels and up regulation of glucose transporter 2-peroxisome proliferator along with activation of gamma receptor, p-insulin receptor substrate1 and Nrf2 in liver and kidney after the administration of AITC. These findings justify the importance of AITC in diabetes prevention. Besides, studies based on diabetes have demonstrated that consumption of SFN (L-SFN) ranging between 112.5 & 225 µM for a few weeks significantly improves markers of type 2 diabetes.107 This was comparable to the study of Waterman108 who investigated the use of ITC rich Moringa oleifera extract as anti-diabetic food in diet induced obese C57BL/6J mice. They observed that in 5% moringa concentrate (MC), the level of blood plasma level of glucose regulators, inflammatory cytokines (IL-1β & TNFα) and cholesterol were lowered down effectively and MCs considerably decreased the glucose production by approximately 60% in H114E liver cells at 10µg/ml and 1µM concentrations, respectively. These observations confirmed the use of moringa as dietary agent that prevents type-2 diabetes by demonstrating that moringa isothiocyanate concentrate caused considerable reduction in weight gain, hepatic adiposity, gluconeogenesis, insulin, cholesterol and inflammatory markers. Apart from these ITCs, SFN too was reported to prevent pathological changes in glomerulus and promoted renal function and blood flow in experimental mouse in which diabetes was induced by using streptozotocin.16 Henceforth, these findings encourage the potential application of ITCs as therapeutic agent against diabetes.
Isothiocyanates as antimicrobial agents
Isothiocyanates have wide range of antimicrobial activity and are generally regarded as safe (GRAS) compounds and hence are allowed to be added to the food.21 For instance, naturally extracted allyl isothiocyanate is already an acceptable preservative in Japan as it has gained GRAS status. These ITCs possess inhibitory activity against food spoilage bacteria species such as Escherichiacoli, Salmonella ssp., Bacillus ssp. etc., but it has been observed that gram-negative bacteria are more sensitive towards isothiocyanates as compared to gram-positive bacteria. The range of antimicrobial spectrum of ITCs is between 0.02 to 200mg/L and aromatic ITCs like benzyl isothiocyanate and phenethyl isothiocyanate are more potent antimicrobial agents than aliphatic ITCS such as allyl isothiocyanates.27,109 Recently, the potency of AITC, BITC & PITC was investigated against Aspergillus parasiticus as it is known to produce aflatoxins (AFs) which are toxic and carcinogenic metabolites found as contaminants in bread, corn, peanut, cottonseed and nuts. It was found that all the three ITCs showed activity against Aspergillus parasiticus and reduced the fungal growth along with 50% decrease in the viability at concentrations ≥0.5mg/L in the liquid medium.110 The antibacterial property of benzyl isothiocyanate (BITC) is owing to its chemical structure as it has both lipophilic as well as electrophilic properties which render it to penetrate through the outer bacterial membrane and disturb the ability of bacteria to maintain cellular integrity and due to its effectiveness it is now being professionally prescribed as an antibiotic to treat infections of respiratory and urinary tracks.23,111 The possible actions of ITCs against microbial activity include reduction of oxygen consumption, depolarization of mitochondrial membrane and intracellular accumulation of reactive oxygen species (ROS) or, inhibition of ATP binding site of P-type ATPase via cystein residue attack in mammals and some bacteria.112,113
Even after many investigations and studies, the proper mechanism of action of ITCs as antimicrobial is unclear, but some action mechanisms may be hypothesized such as, a) ITCs may inhibit functioning of enzymes or accumulate in bacteria as small thiols or thioredoxin dithiocarbamate conjugates and alter the structure of proteins, b) by binding to thiol or amine groups, ITCs may alter the specialized enzymic activities and halt genes transcription, and c) it can cause the oxidative stress which may initiate some oxidation-reduction mechanism that can affect the microbial growth and inhibition.27
Table 2: Health benefits of isothiocyanates.
| Effect | Component | Factor/ Micro-organisms affected | Mechanism | Reference |
| Neuroprotection | Glucomoringin(GMG) 6(Methylsulfinyl)hexyl isothiocyanate (6- SITC)Allyl isothiocyanate (AITC) |
Nrf2 and ARE proteasome catalytic subunit 26SERK1/2 |
Rescission of oxidative stress-induced neurogeneration via Nrf2 and and activation of antioxidant response element -MSITC amended Aβ1-42O-induced memory losses. Upregulation of proteasome catalytic subunit 26S. Rise in Glutathione (GSH) level by activating the nuclear Nrf2 and causing resistance to neuronal apoptosisReduction in H2O2-induced blood cerebrospinal fluid barrier disruption as well as epithelial cell death |
114 115
116 |
| Anti-diabetic | AITC SFNMoringa extract-ITCs |
PPAR-γ and IRS-1p NRF2Increased levels of IRS-1p, IRS-1, IRS-2, Akt1 and GLUT4 | Decrease in blood glucose, total cholesterol, triglycerides and creatinine levels and up regulation of glucose transporter 2-peroxisome proliferator along with activation of gamma receptor, p-insulin receptor substrate1 and Nrf2 in liver and kidneyPrevention of pathological changes in glomerulus and promoted renal function and blood flow. Decreases glucose production from hepatic cells by activation of nuclear translocation of nuclear factor erythroid 2-related factor 2 (NRF2) and decreased expression of key enzymes in gluconeogenesisHigher levels of proteins involved in insulin signaling. Reduction in weight gain, hepatic adiposity, gluconeogenesis, insulin, cholesterol and inflammatory markers | 106
107
108 |
| Antimicrobial | AITC, BITC & PITC | Aspergillus parasiticus H. pylori | Reduction of oxygen consumption, depolarization of mitochondrial membrane and intracellular accumulation of reactive oxygen species (ROS) or, inhibition of ATP binding site of P-type ATPase via cystein residue attackH. pylori gastric infection occurs by its survival in the low pH of the stomach due to urease production which converts urea into ammonia for neutralization of the acidic environment of the stomach. SFN is able to cause H. pylori urease inactivation and hence, its bactericidal activity | 110
109 |
| Cardio-protection | 4-Carboxyphenyl isothiocyanate (4-CPI)SFN |
K-ATP channelAKT/Protein Kinase B and ERK 1/2 | Induce vaso-relaxation effects on coronary artery along with membrane hyper polarization on vascular smooth muscles due to the activity of 4-CPI as H2S donor which helps in activation of mitochondrial K+-ATP channelActivation of AKT/Protein Kinase B and extracellular signal regulated Kinase 1 and 2 (ERK 1/2) signaling pathway that are important for cardiac cell survival and inhibition of phase I enzymes, angiogenesis and inflammation | 104
107 |
| Cancer prevention | SFNITCs | NF-κBARE/EpRE | Activation of phase II enzymes and suppression of the phosphorylation of the inhibitory proteins 1κBα and 1κB Kinase to restrain the nuclear translocation of NF-κB and prevent inflammationITCs promote phase II enzymes (quinone reductase and glutathione S-transferase) by upregulation by the antioxidant or electrophile response element (ARE/EpRE) | 89
90 |
| Antioxidant | ErucinSFN | AREKEAP1/Nrf2/ARE | Possess hydro peroxide scavenging activity and induce enzymes like thioredoxin reductase1 (TrxR1) that act as cellular antioxidantsActivation of several antioxidant enzymes (AO) that help in maintaining redox potential and activity of free radical scavengers such as vitamins A, C and E through KEAP1/Nrf2/ARE pathway | 9218 |
Application of Isothiocyanates in Foods
ITCs in food packaging
The antimicrobial and health-promoting properties make ITCs a good candidate for the application in food packaging, therefore, several authors have investigated their utilization in active packaging. There are many reports dating back to twentieth century that support the use of vaporized allyl isothiocyanate (AITC) for the preservation of various products such as raw beef, egg sandwich, noodles, ground pork, sliced raw tuna, pasta and pears.117 Winther and Nielsen118 investigated the applicability of allyl isothiocyanate in active packaging of cheese. They reported that AITC could increase the storage stability of cheese from 18 weeks (control) to 28 weeks on combination with modified atmosphere (MA) packaging, and, 4.5 (control) weeks to 28 weeks when combined with atmospheric air (AA). Meanwhile, the flavor profile of cheese in MA packaging with AITC changed dramatically whereas, in AA packaging with AITC, CO2 was not used therefore less oxidation occurred and it had a better fresh taste. According to their investigation, AITC extended the shelf life because it effectively inhibited the activity of intracellular enzymes present in Penicillium nalgiovense & Debaryomyces hansenii and suppressed their growth in cheese. Thus, it was inferred that packaging of cheese with AITC and atmospheric air can be a good substitute for modified atmosphere packaging of cheese. Nowadays, the demand for ecofriendly packaging has been escalated, so scientists have developed Polylactic acid (PLA) film to form fibers for food packaging. Polylactic acid is a synthetic polyester that is synthesized either from lactic acid or its dimer lactide. These are suitable for food packaging as these possess strong mechanical properties, are non toxic and environmental friendly. Kara119 used PLA film to encapsulate allyl isothiocyanate (AITC) and inferred that the discharge of allyl isothiocyanate from fiber was temperature dependent and below 4oC temperature, the encapsulated allyl isothiocyanate remained within fibers without losing activity and even showed stability for weeks at room temperature. When the fiber was tested against bacteria Listeria innocua and Escherichia coli, it exhibited the active inhibition of their growth in the food product.
ITCs can also be used as antimicrobial coating on bottles to effectively halt the growth of Salmonella in liquid egg albumen (egg white) and prevent human Salmonellosis disease. The coating of glass jar consists of mixture of polylactic acid polymer and antimicrobial components such as 100-500 µl allyl isothiocyanate (AITC), 250 mg nisin and 250 mg zinc oxide nanoparticles. This coating was reported to completely inactivate 3-7 log CFU ml-1 of Salmonella after 7 and 21 days of storage, respectively, therefore, proving effective against Salmonella sp.120 The effectiveness of AITC as antimicrobial coating against Salmonella was further confirmed by Chen121 as they used AITC in combination with chitosan coating and nisin on whole fresh cantaloupes. They observed that on increasing the concentration of AITC (10 to60 μl/ml), the antibacterial activity of coating against Salmonella was also improved. The 60 μl/ml AITC in chitosan coating effectively decreased more than 5 log10 CFU/cm2 of Salmonella and totally inactivated the growth of yeasts and molds up to 14 days of storage at 10oC. Therefore, these results further add to the possibility of utilizing AITC as an antimicrobial coating for the packaging of foods.
Likewise, ITCs can be microencapsulated and then added to food products in order to provide antibacterial effect. AITC can be encapsulated in Halloysite nanotubes (HNTs) to add to coating material made up of sodium polyacrylate. These are made up of natural aluminosilicate clay and have a hollow nanotubular morphology which provides a large surface area-to-volume ratio and high porosity. Halloysite nanotubes are non-toxic, good thermal stability and biocompatibility therefore can act as a good food packaging material. This complex coating material is effective against E. coli at concentration of 25 μg/mL and inhibits about 96% at 200µg/mL but is less effective against S. aureus when used in concentration between 25-200 µg/mL thus, it can be applied as packaging material for foods.122
ITCs in improving shelf life of minimally processed fruits and vegetables
ITCs can be used for enhancing the shelf life of fermented foodstuffs such as tofu and kimchi, and of minimally processed fruits e.g. strawberries, apples and blueberries. Banerjee123 applied AITC for extending the shelf life of minimally processed shredded cabbage and inhibit browning. They found that AITC inhibited the growth of microflora in concentration dependent manner and treatment of samples with 0.05µL/mL and 0.1µL/mL AITC reduced the mesophilic count to acceptable limit (˃107cfu/g) for upto 12 days at 10oC along with preventing browning and retaining aroma of good quality upto 10 days. The possible underlying mechanism of this preservation might be due to the property of AITC to generate ROS such as H2O2 during storage that damages the membrane and DNA of microbes resulting in bactericidal activity. These observations led to the conclusion that AITC can prove to be a beneficial compound for preservation of fresh cut vegetables. Later, AITC was also investigated for its potency in enhancing the nutritional quality of Kiwifruit and it was observed that a 5 h treatment with 0.15 mg/L AITC combined with CA (controlled atmosphere) reduced the loss of ascorbic acid content as well as total polyphenol content in kiwifruit.124
Application of AITC as preservative of fruits and vegetables is paradoxical as it has strong pungent odor which affects the aromatic appeal of the product, therefore, scientists have suggested the method of its microencapsulation in gelatin, gum arabic or chitosan.125 For instance, addition of (less than 0.1%) microencapsulated AITC in gum arabic increased the pH and texture of Kimchi while decreasing the number of Leuconostoc and Lactobacillus species thereby effectively enhanced the shelf life without affecting its nutritional quality.126 Later on, Wu125 microencapsulated AITC in a concentration range of 0.5-1.0 g/L with gum arabic and gelatin to preserve mature green tomatoes as it prevented degradation by maintaining the acidity of tomatoes.
Fruit juices are considered as health drinks since ancient time and regular fortification has been applied to increase their neutraceutical properties. Diatuo127 used cauliflower byproducts (CBP) as source of ITC and added this ITC concentrate (10-40%) into apple juice. The results obtained showed that apple beverage with 10% ITC extract well preserved the sensorial properties along with color in comparison to control sample.
ITCs as antimicrobial agents in foods
Due to the property of microbial inhibition, ITCs are already utilized in various forms to prevent microbial growth in food. ITCs such as AITC, BITC and PITC can be used as preservative in bakery products to inhibit development of toxic compounds like aflatoxins. In a study, it was reported that at a concentration of 5µL/L, the shelf life of bread loaf treated with AITC increased from 4 (control) to 8 days due to inhibition of fungal growth.106 This was further confirmed by investigation of Torrijos128 in which they used oriental mustard flour (OMF) that contains AITC as principal compound, to develop a bioactive sauce with antifungal properties for extending the shelf life of Pita bread. The result of their study demonstrated a shelf life increase of 2 and 3 days with employment of 33 & 50 mg/g of OMF. The effectiveness of AITC as a preservative was further confirmed by Lopes129 as they evaluated antifungal activity of AITC against Penicillium nordicumin vitro and in frozen pizzas. They noticed that even the lower doses (0.125µL/L) reduced the mycelia growth upto 66% so the higher doses successfully suppressed visual growth for 7 days as well as inhibited colonies of P. nordicum in pizza stored in refrigerated conditions and doubled the shelf life at the concentration of 25 µL AITC/L.
Ethyl isothiocyanate together with allyl isothiocyanate is used as an antifungal agent against infection of Botrytis cinerea and Penicillium expansum on apples and phenyl isothiocyanate (PITC) possesses antifungal activity against Aspergillus niger, Penicillium cyclopium and Rhizopus oryzae.130 Whereas, allyl isothiocyanate is a potent insect attractant that has high antimicrobial activity in liquid media and can destroy bacterial as well as fungal pathogens on fresh produce, bread, meat and cheese.131 It is used as preservative in the food industry since late 20th century in Japan due to its dual role, as a flavor compound and microbicidal agent.132 Various mechanisms have reported that when AITC is used in the form of vapor, it can damage the cell wall integrity and lead to the leakage of cellular metabolite. Some suggested that AITC when exposed to bacterial cells resulted in creation of pores on cell membranes which caused leakage of intracellular substances.133
Lactoperoxidase (LPO) in preventing bacterial growth in dairy products
Lactoperoxidases (LPO) reduce the growth of food borne pathogens in several food products and hence increase their shelf life due to its bactericidal and bacteriostatic properties.134 It is composed of lactoperoxidase enzyme, thiocyanate and hydrogen peroxide. This system produces hypothiocyanite which shows bactericidal properties against gram-positive bacteria as well as gram-negative bacteria. It can be used to improve the shelf life of various foods like fruits, chicken meat and cheese. Al-Baarr135 studied the use of lactoperoxidases for reducing pathogenic bacteria in milk and other dairy products. They observed that LPO significantly decreased the population of E. coli. The possible reason for its inhibitory effect on bacteria is the presence of OSCN¯ compounds in LPOs that destroy bacteria by disrupting sulfhydryl group (-SH) on proteins from bacterial cytoplasmic membrane.136 They further reported that reduction in the amount of lactose might improve the action of LPOs as they inhibit the activity of LPO by hindering its enzymatic activity. Thus, they concluded that lactoperoxidases has moderate antibacterial effect on E. coli in whole milk, skim milk and whey and hence can efficiently be used as antibacterial reagent in reduced-lactose dairy products.
Summary and Conclusion
Isothiocyanates are one of those promising compounds that contain many health benefits including chemoprevention, neuro-protection, antioxidant activity, anti-obesity, cardio-protective, fungicidal and antibacterial. They activate various important channels and pathways like Nrf2/ERK and CDK-cyclin complex along with regulating phase I and phase II enzymes and down-regulating NFκB factor to prevent inflammation of body tissues. Such mechanisms make them effective in inducing apoptosis, inhibition of cell cycle progression and angiogenesis. Further research on the properties and mechanisms of action of isothiocyanates in disease prevention will provide a promising future in finding a cure to these disorders. Apart from this, isothiocyanates have proven to be effective against harmful bacteria and fungi. Their power packed properties provide the prospects of using them in various food applications like bio-fibers, active packaging to increase the shelf life of foodstuffs, and as natural food additives such as antioxidants and antimicrobial agents. The non toxicity of most of isothiocyanates adds to their properties and overall acceptability. Great efforts have been made to improve the stability and to keep the characteristic taste and flavour profile of ITCs intact. Still, there are various mechanisms of isothiocyanates that are yet to be studied. Thus, more detailed studies are required to explore properties of ITCs for their potential use and acceptability in food as well as other industries.
Acknowledgement
The authors want to thank to the Department of Food Technology, Maharishi Dayanand University,Rohtak, Haryana, India for providing assistance.
Conflict of Interest
The authors declare that there is no conflict of interest.
Funding Sources
This work did not receive any specific grant from funding agencies.
References
- Brown K. K., M. B. Hampton, Biological targets of isothiocyanates, et. biophysica. Acta., 2011, 1810, 888–894. https://doi.org/10.1016/j.bbagen.2011.06.004.
CrossRef - Bell L., Oloyede O. O., Lignou S., Wagstaff C., Methven L. Taste and flavor perceptions of glucosinolates, isothiocyanates, and related compounds, Nutr. Food Res., 2018, 62, e1700990. https://doi.org/10.1002/mnfr.201700990.
CrossRef - Kala C., Ali S., Ahmad N., Jamal Gilani S., Ali Khan N. Isothiocyanates: a review, J. Pharmacogn., 2018, 5, 71-89. https://doi.org/10.22127/rjp.2018.58511.
CrossRef - Jang M., Hong E., Kim G. H. Evaluation of antibacterial activity of 3-butenyl, 4-pentenyl, 2-phenylethyl, and benzyl isothiocyanate in Brassica vegetables, Food Sci., 2010, 75, 412–416. https://doi.org/10.1111/j.1750-3841.2010.01725.x.
CrossRef - Hayes J. D., Kelleher, M. O., Eggleston I. M. The cancer chemo preventive actions of phytochemicals derived from glucosinolates, J. Nutr., 2008, 47, 73–88. https://doi.org/10.1007/s00394-008-2009-8.
CrossRef - Giacoppo S., Galuppo M., Montaut S., Iori R., Rollin P., Bramanti P., Mazzon E. An overview on neuroprotective effects of isothiocyanates for the treatment of neurodegenerative diseases, Fitoterapia, 2015, 106, 12-21. https://doi.org/10.1016/j.fitote.2015.08.001.
CrossRef - Sotelo T., Lema M., Soengas P., Cartea M. E., Velasco P. In Vitro activity of glucosinolates and their degradation products against brassica-pathogenic bacteria and fungai, Environ. Microbiol., 2014, 81, 432-440. https://doi.org/10.1128/AEM.03142-14.
CrossRef - Guerrero-Díaz M. M., Lacasa-Martínez C. M., Hernández-Piñera A., Martínez-Alarcón V. Lacasa-Plasencia A. Evaluation of repeated biodisinfestation using Brassicacarinata pellets to control Meloidogyneincognita in protected pepper crops, J. Agric. Res., 2013, 11, 485-493. https://doi.org/10.5425/sjar/2013112-3275.
CrossRef - Sarwar M., Kirkegaard J. A., Wong P. T. W., Desmarchelier J. M. Bio fumigation potential of brassicans, Plant and Soil, 1998, 201, 103-112. https://doi.org/10.1023/A:1004381129991.
CrossRef - Kuchernig J. C., Burow M. Wittstock U. Evolution of specifier proteins in glucosinolates containing plants, BMC Evol. Biol., 2012, 12, 127. https://doi.org/10.1186/1471-2148-12-127.
CrossRef - Matusheski N. V., Jeffery E. H. Comparison of the bioactivity of two glucoraphanin hydrolysis products found in broccoli, sulforaphane and sulforaphane nitrile, Agric. Food Chem., 2001, 49, 5743–5749. https://doi.org/10.1021/jf010809a.
CrossRef - Matusheski N. V., Juvik J. A., Jeffery E. H. Heating decreases epithiospecifier protein activity and increases sulforaphane formation in broccoli, , 2004, 65, 1273–1281. https://doi.org/10.1016/j.phytochem.2004.04.013.
CrossRef - Bricker G. V., Riedl K. M., Ralston R. A. Tober K. L. Oberyszyn T. M., Schwartz S. J. Isothiocyanate metabolism, distribution and interconversion in mice following consumption of thermally processed broccoli sprouts or purified sulforaphane, Nutr. Food Res.,2014, 58, 1991–2000. https://doi.org/10.1002/mnfr.201400104.
CrossRef - Munday R., Munday C. M. Induction of phase II detoxification enzymes in rats by plant-derived isothiocyanates: comparison of allyl isothiocyanate with sulforaphane and related compounds, Agric. Food Chem., 2004, 52, 1867-1871. https://doi.org/10.1021/jf030549s.
CrossRef - Tumer T. B., Rojas-Silva P., Poulev A., Raskin I., Waterman C. Direct and indirect antioxidant activity of polyphenol- and isothiocyanate-enriched fractions from Moringaoleifera, Agric. Food Chem., 2015, 63, 1505–1513. https://doi.org/10.1021/jf505014n.
CrossRef - Dinkova-Kostova A.T., Kostov R. V. Glucosinolates and isothiocyanates in health and disease, Trends Mol. Med., 2012, 18, 337-347. https://doi.org/10.1016/j.molmed.2012.04.003.
CrossRef - Traka M., Mithen R. Glucosinolates, Isothiocyanates and human health, Rev., 2009, 8, 269-282. https://doi.org/10.1007/s11101-008-9103-7.
CrossRef - Boddupalli R., Mein J. R., Lakkanna S. James D. R. Induction of phase 2 antioxidant enzymes by broccoli sulforaphane: perspectives in maintaining the antioxidant activity of vitamins A, C and E, Genet., 2012, 3, 7. https://doi.org/10.3389/fgene.2012.00007.
CrossRef - Srivastava S. K., Singh S. V. Cell cycle arrest, apoptosis induction and inhibition of nuclear factor kappa B activation in anti-proliferative activity of benzyl isothiocyanate against human pancreatic cancer cells, Carcinogenesis, 2004, 25, 1701-1709. https://doi.org/10.1093/carcin/bgh179.
CrossRef - Kaiser S. J., Mutters N. T., Blessing B., Günther F. Natural isothiocyanates express antimicrobial activity against developing and mature biofilms of Pseudomonasaeruginosa, Fitoterapia, 2017, 119, 57–63. https://doi.org/10.1016/j.fitote.2017.04.006.
CrossRef - Romeo L., Iori R., Rollin P., Bramanti P., Mazzon E. Isothiocyanates: An overview of their antimicrobial activity against human infections, Molecules,2018, 23, 624. https://doi.org/10.3390/molecules23030624.
CrossRef - Aires A., Mota V. R., Mota, M. J., Saavedra E., Rosa A. S. Bennett R. N. The antimicrobial effects of glucosinolates and their respective enzymatic hydrolysis products on bacteria isolated from the human intestinal tract, Appl. Microbiol., 2009, 106, 2086–2095. https://doi.org/10.1111/j.1365-2672.2009.04180.x.
CrossRef - Sofrata A., Santangelo E. M., Azeem M., Borg-Karlson A. K., Gustafsson A., Pütsep K. Benzyl isothiocyanate, a major component from the roots of Salvadorapersica is highly active against Gram-negative bacteria, PLOS One, 2011, 6, e23045. https://doi.org/10.1371/journal.pone.0023045.
CrossRef - Wilson A. E., Bergaentzlé M., Bindler F., Marchioni E., Lintz, A., Ennahar S. In vitro efficacies of various isothiocyanates from cruciferous vegetables as antimicrobial agents against foodborne pathogens and spoilage bacteria, Food Cont., 2013, 30, 318-324. https://doi.org/10.1016/j.foodcont.2012.07.031.
CrossRef - Smolinska U., Morra M. J., Knudsen G. R., James R. L. Isothiocyanates produced by Brassicaceae species as inhibitors of Fusarium oxysporum, Plant Disease, 2003, 87, 407- 412. https://doi.org/10.1094/PDIS.2003.87.4.407.
CrossRef - Cejpek K., Valusek J., Velisek J. Reactions of Allyl Isothiocyanate with alanine, glycine and several peptides in model systems, Agric. Food Chem., 2000, 48, 3560- 3565. https://doi.org/10.1021/jf991019s.
CrossRef - Dufour V., Stahl M., Baysse C. The antibacterial properties of isothiocyanates, Microbiology, 2015, 161, 229–243. https://doi.org/10.1099/mic.0.082362-0.
CrossRef - Blažević I., Mastelić J. Glucosinolate degradation products and other bound and free volatiles in the leaves and roots of radish (Raphanussativus), Food Chem., 2009, 113, 96-102, https://doi.org/10.1016/j.foodchem.2008.07.029.
CrossRef - Zhenxin G., Qianghui G., Yingjuan G. Factors influencing glucoraphanin and sulforaphane formation in brassica plants: A review. J. Integr. Agric., 2012, 11, 1804–1816. https://doi.org/10.1016/S2095-3119(12)60185-3.
CrossRef - Zinoviadou K. G., Galankis C. M. Glucosinolates and respective derivatives (isothiocyanates) from plants. In: Puri M. (eds) Food Bioactives, Springer, Cham., 2017, 2017, 3-22. https://doi.org/10.1007/978-3-319-51639-41.
CrossRef - Śmiechowska A., Bartoszek A., Namieśnik J. Determination of glucosinolates and their decomposition products-indoles and isothiocyanates in cruciferous vegetables, crit. rev. anal. chem., 2010, 40, 202-216. https://doi.org/10.1080/10408347.2010.490489.
CrossRef - Nakamura T., Murata Y., Nakamura Y. Characterization of benzyl isothiocyanate extracted from mashed green papaya by distillation, Food Chem., 2019, 299, 125118. https://doi.org/10.1016/j.foodchem.2019.125118.
CrossRef - Arora R., Singh B., Vig A. P., Arora S. Conventional and modified Hydrodistillation method for the extraction of glucosinolate hydrolytic products: a comparative account, Springer Plus, 2016, 5, 479. https://doi.org/10.1186/s40064-016-2021-z.
CrossRef - Abdel-Kader M. S., Alam P., Kamal Y. T., Alkharfy K. M., Foudah, A. I., Alqasoumi S. I. Optimization of the extraction condition for benzyl isothiocyanate contents in Salvadorapersica roots “Siwak”, J., 2019, 27, 753-755. https://doi.org/10.1016/j.jsps.2019.04.011.
CrossRef - Doheny-Adams T., Redeker K., Kittipol V., Bancroft I., Hartley S. E. Development of an efficient glucosinolate extraction method, Plant Methods, 2017, 13, 17. https://doi.org/10.1186/s13007-017-0164-8.
CrossRef - Ares A. M., Bernal J., Martín M. T., Bernal J. L, Nozal M. J. Optimized formation, extraction, and determination of sulforaphane in broccoli by liquid chromatography with diode array detection, Food Anal. Methods, 2014, 7, 730-740. https://doi.org/10.1007/s12161-013-9766-6.
CrossRef - Hebert M., Mhemdi H., Vorobiev E. Selective and eco-friendly recovery of glucosinolates from mustard seeds (Brassica juncea) using process optimization and innovative pretreatment (high voltage electrical discharges), Food Bioprod. Process., 2020, 124, 11-23. https://doi.org/10.1016/j.fbp.2020.04.009.
CrossRef - Powell E. E., Hill G. A., Juurlink B., Carrier D. J. Glucoraphanin extraction from Cardariadraba: Part 1. Optimization of batch extraction, Chem. Technol. Biotechnol., 2005, 80, 985 – 991. https://doi.org/10.1002/jctb.1273.
CrossRef - Frandsen H. B., Markedal K. E., Belloso O. M., Vega R. S., Fortuny R. S., Sørensen H., Sørensen S., Sørensen J. Effects of novel processing techniques on glucosinolates and membrane associated myrosinases. In Broccoli, J. Food Nutr. Sci., 2014, 64, 17-25. https://doi.org/10.2478/pjfns-2013-0005.
CrossRef - Wang T., Liang H., Yuan Q. Optimization of ultrasonic-stimulated solvent extraction of sinigrin from indian mustard seed (BrassicaJuncea) using response surface methodology, Phytochemical Anal., 2011, 22, 205-213. https://doi.org/10.1002/pca.1266.
CrossRef - Tanongkankit Y., Sablani S. S., Chiewchan N., Devahastin S. Microwave-assisted extraction of sulforaphane from white cabbages: Effects of extraction condition, solvent and sample pretreatment, Food Eng., 2013, 117, 151-157. https://doi.org/10.1016/j.jfoodeng.2013.02.011.
CrossRef - Jaiswal A. K., Abu-Ghannam N. Fermentation-assisted extraction of isothiocyanates from brassica vegetable using box-behnken experimental design, Foods, 2016, 5, 75. https://doi.org/10.3390/foods5040075.
CrossRef - Koo S. Y., Cha K. H., Song D., Lee D., Pan, C. Increased Sulforaphane concentration in Brussels sprout following high hydrostatic pressure treatment, Korean Soc. Appl. Biol. Chem., 2012, 55, 685–687. https://doi.org/10.1007/s13765-012-2123-4.
CrossRef - Westphal A., Riedl K. M., Cooperstone J. L., Kamat S., Balasubramaniam V. M., Schwartz S. J., Böhm V. High-pressure processing of broccoli sprouts: influence on bioactivation of glucosinolates to isothiocyanates, Agric. Food Chem., 2017, 65, 8578–8585. https://doi.org/10.1021/acs.jafc.7b01380.
CrossRef - Ameer K., Shahbaz H. M., Kwon J. Green extraction methods for polyphenols from plant matrices and their byproducts: a review: Polyphenols extraction by green methods, Rev. Food Saf., 2017, 16, 295-315. https://doi.org/10.1111/1541-4337.12253.
CrossRef - Li L., Lee W., Lee W. J., Auh J. H., Kim S. S, Yoon J. Extraction of allyl isothiocyanate from wasabi (WasabiajaponicaMatsum) using supercritical carbon dioxide, Food Sci. Biotechnol., 2010, 19, 405–410. https://doi.org/10.1007/s10068-010-0057-3.
CrossRef - Aguiló-Aguayo I., Suarez M., Plaza L., Hossain M. B., Brunton N., Lyng J. G., Rai D. K. Optimization of pulsed electric field pre-treatments to enhance health-promoting glucosinolates in broccoli flowers and stalk, Sci. Food Agri., 2015, 95, 1868–1875. https://doi.org/10.1002/jsfa.6891.
CrossRef - Banožic M., Jozinovic A., Grgic J., Milicevic B., Jokic S. High voltage electric discharge for recovery of chlorogenic acid from tobacco waste. sustainability, 2021, 13, 4481. https://doi.org/10.3390/ su13084481
CrossRef - Barba F.J., Boussetta N., Vorobiev E. Emerging technologies for the recovery of isothiocyanates, protein and phenolic compounds from rapeseed and rapeseed press-cake: Effect of high voltage electrical discharges. Food Sci. Emerg. Technol., 2015, 31, 67-72. https://doi.org/10.1016/j.ifset.2015.06.008
CrossRef - Rodrigues L., Silva I., Poejo J., Serra A.T., Matias A.A., Simplício A.L., Bronze M.R., Duarte C.M.M. Recovery of antioxidant and antiproliferative compounds from watercress using pressurized fluid extraction. RSC Adv. 2016, 6, 30905–30918. https://doi.org/10.1039/C5RA28068K
CrossRef - Luang-In V., Rossiter J. Stability studies of isothiocyanates and nitriles in aqueous media, Songklanakarin J. Sci. Technol., 2015, 37, 625-630. https://rdo.psu.ac.th/sjstweb/journal/37-6/37-6-3.pdf
- Liu T. T., Yang T. S. Stability and antimicrobial activity of allyl isothiocyanate during long-term storage in an oil-in-water emulsion, Food Sci., 2010, 75, 445–451. https://doi.org/10.1111/j.1750-3841.2010.01645.x.
CrossRef - Cejpek, Urban J., Velíšek J., Hrabcová H. Effect of sulphite treatment on allyl isothiocyanate in mustard paste, Food Chem., 1998, 62, 53-57. https://doi.org/10.1016/S0308-8146(97)00181-7.
CrossRef - Van Eylen D., Oey I., Hendrickx M., Van Loey A. Kinetics of the stability of broccoli (Brassica oleracea Italica) myrosinase and isothiocyanates in broccoli juice during pressure/temperature treatments, J. Agric. Food Chem.,2007, 55, 2163–2170. https://doi.org/10.1021/jf062630b.
CrossRef - Fang Z., Bhandari B. Encapsulation of polyphenols – a review, Trends Food Sci. Technol., 2010, 21, 510-523. https://doi.org/10.1016/j.tifs.2010.08.003.
CrossRef - Wu H., Yan X., Liang H., Yuan Q. Preparation and stability investigation of the inclusion complex of sulforaphane with hydroxypropyl-cyclodextrin, Polym., 2010, 82, 613-617. http://doi.org/10.1016/j.carbpol.2010.05.020.
CrossRef - Neoh L., Yamamoto C., Ikefuji S., Furuta T., Yoshii H. Heat stability of allyl isothiocyanate and phenyl isothiocyanate complexed with randomly methylated β-cyclodextrin, Food Chem., 2012, 131, 1123-1131. https://doi.org/10.1016/j.foodchem.2011.09.077.
CrossRef - Heaney R. P. Factors influencing the measurement of bioavailability, taking calcium as a model, Nutr., 2001, 131, 1344–1348. https://doi.org/10.1093/jn/131.4.1344S.
CrossRef - Wood R. J. In: Caballero B, Allen L, Prentice A (eds) Encyclopedia of human nutrition. Elsevier Ltd, Oxford, UK, 2005. https://www.academia.edu/44913390/Encyclopedia_of_Human_Nutrition_by_Benjamin_Caballero_ Lindsay_Allen_Andrew_Prentice
- Oliviero T., Verkerk R., Dekker M. Isothiocyanates from brassica vegetables-effects of processing, cooking, mastication, and digestion, Nutr. Food Res.,2018, 62, 1701069. https://doi.org/10.1002/mnfr.201701069.
CrossRef - Shapiro T. A., Fahey J. W., Dinkova-Kostova A. T., Holtzclaw W. D., Stephenson K. K., Wade K. L., Ye L., Talalay P. Safety, tolerance, and metabolism of broccoli sprout glucosinolates and isothiocyanates: A clinical phase I study, Cancer-an Int. J., 2006, 55, 53–62. https://doi.org/10.1207/s15327914nc5501_7.
CrossRef - Barba F. S., Nikmaram N., Rooinejad S., Khelfa A., Zhu Z., Koubaa M. Bioavailability of glucosinolates and their breakdown products: impact of processing, Front. Nutr., 2016, 3, 24. https://doi.org/10.3389/fnut.2016.00024.
CrossRef - Verkerk R., Van der Gaag M. S., Dekker M., Jongen W. M. Effects of processing conditions on glucosinolates in cruciferous vegetables, Cancer letters, 1997, 114, 193-194. https://doi.org/10.1016/s0304-3835(97)04661-2.
CrossRef - McNaughton S., Marks G. Development of a food composition database for the estimation of dietary intakes of glucosinolates, the biologically active constituents of cruciferous vegetables, J. Nutr., 2003, 90, 687-697. https://doi.org/10.1079/BJN2003917.
CrossRef - Getahun S. M., Chung F. L. Conversion of glucosinolates to isothiocyanates in humans after ingestion of cooked watercress, Cancer Epidemiol. Biomarkers Prev., 1999, 8, 447–451. PMID: 10350441.
- Krul C., Humblot C., Philippe C., Vermeulen M., van Nuenen M., Havenaar R., Rabot S. Metabolism of sinigrin (2-propenyl glucosinolate) by the human colonic microflora in a dynamic in vitro large-intestinal model, Carcinogenesis, 2002, 23, 1009-1016. https://doi.org/10.1093/carcin/23.6.1009.
CrossRef - Jones R. B., Frisina C. L., Winkler S., Imsic M., Tomkins R. B. Cooking method significantly effects glucosinolate content and sulforaphane production in broccoli florets, Food Chem., 2010, 123, 237–242. https://doi.org/10.1016/j.foodchem.2010.04.016.
CrossRef - Miglio C., Chiavaro E., Visconti A., Fogliano V., Pellegrini N. Effects of different cooking methods on nutritional and physicochemical characteristics of selected vegetables, Agric. Food Chem., 2008, 56, 139-147. https://doi.org/10.1021/jf072304b.
CrossRef - Rungapamestry V., Duncan A., Fuller Z., Ratcliffe B. Influence of blanching and freezing broccoli (Brassica oleracea italica) prior to storage and cooking on glucosinolate concentrations and myrosinase activity, Eur. Food Res. Technol., 2008, 227, 37-44. https://doi.org/10.1007/s00217-007-0690-0.
CrossRef - Song L., Thornalley P. J. Effect of storage, processing and cooking on glucosinolate content of Brassica vegetables, Food Chem. Toxicol., 2007, 45, 216-224. https://doi.org/10.1016/j.fct.2006.07.021.
CrossRef - Nugrahedi P. Y., Oliviero T., Heising J. K., Dekker M., Verkerk R. Stir-frying of Chinese cabbage and pakchoi retains health-promoting glucosinolates, Plant Foods Hum. Nutr., 2017, 72, 439-444. https://doi.org/10.1007/s11130-017-0646-x.
CrossRef - Martinez-Villaluenga C., Peñas E., Frias J., Ciska E., Honke J., Piskula M. K., Kozlowska H., Vidal-Valverde C. Influence of fermentation conditions on glucosinolates, ascorbigen, and ascorbic acid content in white cabbage (Brassica oleracea capitata cv. Taler) cultivated in different seasons, J. Food Sci., 2009, 74, 62–67. https://doi.org/10.1111/j.1750-3841.2008.01017.x.
CrossRef - Suzuki C., Ohnishi-Kameyama M., Sasaki K., Murata T., Yoshida M. Behavior of glucosinolates in pickling cruciferous vegetables, Agric. Food Chem., 2006, 54, 9430-9436. https://doi.org/10.1021/jf061789l.
CrossRef - Rangkadilok N., Tomkins B., Nicolas M.E., Premier R. R., Bennett R. N., Eagling D. R., Taylor P. W. J. The effect of post-harvest and packaging treatments on Glucoraphanin concentration in broccoli (Brassica oleracea italica), J. Agric. Food Chem., 2002, 50, 7386–7391. https://doi.org/10.1021/jf0203592.
CrossRef - Rodrigues A. S., Rosa E. A. S. Effect of post‐harvest treatments on the level of glucosinolates in broccoli. Sci. Food Agric., 1999, 79, 1028-1032. https://doi.org/10.1002/(SICI)1097-0010(19990515)79:7%3C1028::AID-JSFA322%3E3.0.CO;2-I.
CrossRef - D’Auria M., Mauriello G., Recioppi R. SPME-GC-MS analysis of horseradish (Armoracia rusticana) [solid-phase microextraction-gas chromatography-mass spectrometry]. J. Food Sci., 2004, 16, 487-490. https://doi.org/10.1111/j.1365-2621.2004.00889.x.
CrossRef - Agneta R., Möllers C., Rivelli A. R. Horseradish (Armoracia rusticana), a neglected medical and condiment species with a relevant glucosinolate profile: a review, Resour. Crop Evol. 2013, 60, 1923–1943. https://doi.org/10.1007/s10722-013-0010-4.
CrossRef - Cavallo C., Cicia G., Del Giudice T., Sacchi R., Vecchio R. Consumers’ perceptions and preferences for bitterness in vegetable foods: the case of extra-virgin olive oil and brassicaceae- a narrative review, Nutrients, 2019, 11, 1164-1184. https://doi.org/10.3390/nu11051164.
CrossRef - Macpherson L. J., Dubin A. E., Evans M. J., Marr F., Schultz P. G., Cravatt B. F., Patapoutian A. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines, Nature, 2007, 445, 541-545. https://doi.org/10.1038/nature05544.
CrossRef - Idikio H. A. Human cancer classification: a systems biology- based model integrating morphology, cancer stem cells, proteomics, and genomics, Cancer, 2011, 2, 107–115. https://doi.org/10.7150/jca.2.107.
CrossRef - Molina Vargas L. F. Mechanism of action of isothiocyanates: A review, AgronomíaColombiana, 2013, 31, 68-75. https://www.researchgate.net/publication/262460316_Mechanism_of_action_of_isothiocyanates_A_review.
- Koene R. J., Prizment A. E., Blaes A., Konety S. H. Shared risk factors in cardiovascular disease and cancer, Circulation, 2016, 133, 1104-1114. https://doi.org/10.1161/CIRCULATIONAHA.115.020406.
CrossRef - Sheweita S. A., Tilmisany A. K. Cancer and phase II drug-metabolizing enzymes, Drug Metab., 2003, 4, 45-58. https://doi.org/10.2174/1389200033336919.
CrossRef - Johnson I. T. Glucosinolates: bioavailability and importance to health, J. Vitam. Nutr. Res.,2002, 72, 26–31. https://doi.org/10.1024/0300-9831.72.1.26.
CrossRef - Steele V. E. Current mechanistic approaches to the chemoprevention of cancer. Biochem. Mol. Biol., 2003, 36, 78–81. https://doi.org/10.5483/bmbrep.2003.36.1.078.
CrossRef - Dorai T., Aggarwal B. B. Role of chemopreventive agents in cancer therapy, Cancer Lett., 2004, 215, 129-140. https://doi.org/10.1016/j.canlet.2004.07.013.
CrossRef - Wattenberg L. W. Inhibition of carcinogenic effects of polycyclic hydrocarbons by benzyl isothiocyanate and related compounds, J. Natl. Cancer Inst. 1977, 58, 395-398. https://doi.org/10.1093/jnci/58.2.395.
CrossRef - Abbaoui B., Lucas C. R., Riedl K. M., Clinton S. K., Mortazavi A. Cruciferous vegetables, isothiocyanates, and bladder cancer prevention, Nutr. Food Res., 2018, 62, 1800079. https://doi.org/10.1002/mnfr.201800079.
CrossRef - Singh A. V. Sulforaphane induces caspase-mediated apoptosis in cultured PC-3 human prostate cancer cells and retards growth of PC-3 xenografts in vivo, Carcinogenesis, 2003, 25, 83- 90. https://doi.org/10.1093/carcin/bgg178.
CrossRef - Yuanfeng W., Gongian X., Jianwei M., Shiwang L., Jun H., Lehe M. Dietary sulphoraphane inhibits histone deacetylases activity in B16 melanoma cells, Funct. Foods, 2015, 18, 182-189. https://doi.org/10.1016/j.jff.2015.07.002.
CrossRef - de Figueiredo S. M., Filho S. A., Nogueira-Machado J. A., Caligiorne R. B. The anti-oxidant properties of isothiocyanates: a review, Recent Pat. Endocr. Metab. Immune drug Discov., 2013, 7, 213–225. https://doi.org/10.2174/18722148113079990011.
CrossRef - Wang W., Wang S., Howie A. F., Beckett G. J., Mithen R., Bao Y. Sulforaphane, Erucin, and iberin up-regulate thioredoxin reductase 1 expression in human MCF-7 cells. Agric. food Chem., 2005, 53, 1417–1421. https://doi.org/10.1021/jf048153j.
CrossRef - Melchini A., Traka M. H. Biological profile of Erucin: A new promising anticancer agent from cruciferous vegetables, Toxins, 2010, 2, 593-612. https://doi.org/10.3390/toxins2040593.
CrossRef - Abbott N. J., Patabendige A. A., Dolman D. E., Yusof S. R., Begley D. J. Structure and function of the blood-brain barrier, Dis., 2010, 37, 13–25. https://doi.org/10.1016/j.nbd.2009.07.030.
CrossRef - Lee L., Sakurai M., Matsuzaki S., Arancio O., Fraser P. SUMO and Alzheimer’s disease, Neuromolecular Med., 2013, 15, 720–736. https://doi.org/10.1007/s12017-013-8257-7.
CrossRef - Tarozzi A., Angeloni C., Malaguti M., Morroni F., Hrelia S., Hrelia P. Sulforaphane as a potential protective phytochemical against neurodegenerative diseases, Med. Cell. Longev., 2013, 2013, 415078. https://doi.org/10.1155/2013/415078.
CrossRef - Brandes M. S., Gray N. E. NRF2 as a therapeutic target in neurodegenerative diseases, ASN neuro, 2020, 12, 1-23. https://doi.org/10.1177/1759091419899782.
CrossRef - Dwivedi S., Rajasekar N., Hanif K., Nath C., Shukla R. Sulforaphane ameliorates okadaic acid-induced memory impairment in rats by activating the nrf2/ho-1 antioxidant pathway, Molecular Neurobiology, 2016, 53, 5310–5323. https://doi.org/10.1007/s12035-015-9451-4.
CrossRef - Dvoriantchikova G., Barakat D., Brambilla R., Agudelo C., Hernandez E., Bethea J. R., Shestopalov V. I., Ivanov D. Inactivation of astroglial NF-kappa B promotes survival of retinal neurons following ischemic injury, J. Neurosci., 2009, 30, 175–185. https://doi.org/10.1111/j.1460-9568.2009.06814.x.
CrossRef - Cristancho A. G., Lazar M. A. Forming functional fat: a growing understanding of adipocyte differentiation, Nature reviews, Mol. Cell Biol., 2011, 12, 722–734. https://doi.org/10.1038/nrm3198.
CrossRef - Suido H., Tanaka T., Tabei T., Takeuchi A., Okita M., Kishimoto T., Kasayama S., Higashino K. A mixed green vegetable and fruit beverage decreased the serum level of low-density lipoprotein cholesterol in hypercholesterolemic patients, Agric. Food Chem., 2002, 50, 3346–3350. https://doi.org/10.1021/jf0116698.
CrossRef - Assad T., Khan R. A., Feroz Z. Evaluation of hypoglycemic and hypolipidemic activity of methanol extract of Brassicaoleracea, J. Nat. Med., 2014, 12, 648–653. https://doi.org/10.1016/S1875-5364(14)60099-6.
CrossRef - Lo C. W., Chen C. S., Chen Y. C., Hsu Y. A., Huang C. C., Chang C. Y., Lin C. J., Lin C. W., Lin H. J., Liu F. T., Wan L. Allyl Isothiocyanate ameliorates obesity by inhibiting galectin-12, Nutr Food Res., 2018, 62, 1700616. https://doi.org/10.1002/mnfr.201700616.
CrossRef - Testai L., Marino A., Piano I., Brancaleone V., Tomita K., Di Cesare Mannelli L., Martelli A., Citi V., Breschi M. C., Levi R., Gargini C., Bucci M., Cirino G., Ghelardini C., Calderone V. The novel H 2 S-donor 4-carboxyphenyl isothiocyanate promotes cardioprotective effects against ischemia/reperfusion injury through activation of mitoK ATP channels and reduction of oxidative stress, Res., 2016, 113, 290–299. https://doi.org/10.1016/j.phrs.2016.09.006.
CrossRef - Lal, Suresh B, Diabetes: causes, symptoms and treatments. In book: Public health environment and social issues in india, edition: 1, Chapter: 5, Publisher: Serials Publications, Editors: B Suresh Lal, 2016, pp.55-67.
- Sahin N., Orhan C., Erten F., Tuzcu M., Defo Deeh P. B., Ozercan I. H., Juturu V., Kazim S. Effects of allyl isothiocyanate on insulin resistance, oxidative stress status, and transcription factors in high-fat diet/streptozotocin-induced type 2 diabetes mellitus in rats, Biochem. Mol. Toxicol., 2019, 33, 22328. https://doi.org/10.1002/jbt.22328.
CrossRef - Axelsson A. S., Tubbs E., Mecham B., Chacko S., Nenonen H. A., Tang Y., Fahey J. W., Derry J., Wollheim C. B., Wierup N., Haymond M. W., Friend S. H., Mulder H., Rosengren A. H. Sulforaphane reduces hepatic glucose production and improves glucose control in patients with type 2 diabetes, Transl. Med., 2017,9, 4477. https://doi.org/10.1126/scitranslmed.aah4477.
CrossRef - Waterman C., Rojas-Silva P., Tumer T. B., Kuhn P., Richard A. J., Wicks S., Stephens J. M., Wang Z., Mynatt R., Cefalu W., Raskin I. Isothiocyanate-rich Moringa oleifera extract reduces weight gain, insulin resistance, and hepatic gluconeogenesis in mice, Nutr. Food Res., 2015, 59, 1013-1024. https://doi.org/10.1002/mnfr.201400679.
CrossRef - Kim M. G., Lee H. S. Growth-inhibiting activities of phenethyl isothiocyanate and its derivatives against intestinal bacteria, J. Food Sci., 2009, 74, 467–471. https://doi.org/10.1111/j.1750-3841.2009.01333.x.
CrossRef - Saladino F., Quiles J. M., Luciano F., Man̋es J., Fernández-Franzón M., Meca G. Shelf life improvement of the loaf bread using allyl, phenyl and benzyl isothiocyanates against Aspergillusparasiticus, LWT – Food Sci. Technol., 2017, 78, 208-214. https://doi.org/10.1016/j.lwt.2016.12.049.
CrossRef - Vig A. P., Rampal G., Thindand T. S., Arora S. Bio- protective effects of glucosinolates – A review, LWT – Food Sci. Technol., 2009, 42, 1561- 1572. https://doi.org/10.1016/j.lwt.2009.05.023.
CrossRef - Calmes B., N’Guyen G., Dumur J., Brisach C. A., Campion C., Iacomi B., Pigné S., Dias E., Macherel D., Guillemette T., Simoneau P. Glucosinolate-derived isothiocyanates impact mitochondrial function in fungal cells and elicit an oxidative stress response necessary for growth recovery, Plant Sci., 2015, 6, 414. https://doi.org/10.3389/fpls.2015.00414.
CrossRef - Breier A., Ziegelhöffer A. “Lysine is the Lord” thought some scientists in regard to the group interacting with fluorescein isothiocyanate in ATP-binding sites of P-type ATPases but, is it not cysteine? Physiol. Biophys., 2000, 19, 253–263.
- Jaafaru M.S., Nordin N., Rosli R., Shaari K., Bako H.Y., Saad N., Noor N.M., Abdull Razis A.F. Neuroprotective effects of glucomoringin-isothiocyanate against H2O2-Induced cytotoxicity in neuroblastoma (SH-SY5Y) cells. Neurotoxicology, 2019, 75, 89-104. https://doi.org/10.1016/j.neuro.2019.09.008
CrossRef - Morroni F., Sita G., Djemil A., D’Amico M., Pruccoli L., Cantelli-Forti G., Hrelia P., Tarozzi A. Comparison of adaptive neuroprotective mechanisms of sulforaphane and its interconversion product erucin in in vitro and in vivo models of parkinson’s disease. Agri. Food Chem., 2018, 66(4), 856-865. https://doi.org/10.1021/acs.jafc.7b04641
CrossRef - Xiang J., Alesi G.N., Zhou N., Keep R.F. Protective effects of isothiocyanates on blood-CSF barrier disruption induced by oxidative stress. J. Physiol. Regul. Integr. Comp. Physiol., 2012, 303(1), R1-7. https://doi.org/10.1152/ajpregu.00518.2011
CrossRef - Kenji I., Keiko T., Risaburo M., Saiko C. Preliminary examination of allyl isothiocyanate vapor for food preservation, Biosci, Biotecnol., Biochem., 1992, 56, 1476-1477. https://doi.org/10.1271/bbb.56.1476.
CrossRef - Winther M., Nielsen P. V. Active packaging of cheese with allyl isothiocyanate, an alternative to modified atmosphere packaging, Food Prot., 2006, 69, 2430–2435. https://doi.org/10.4315/0362-028x-69.10.2430.
CrossRef - Kara H. H., Xiao F., Sarker M., Jin T. Z., Sousa A. M. M., Liu C., Tomasula P., Liu L. Antibacterial poly (lactic acid) (PLA) films grafted with electrospun PLA/allyl isothiocyanate fibers food packaging, Appl. Polym. Sci., 2016, 133, 42475. https://doi.org/10.1002/app.42475.
CrossRef - Jin T., Gurtler J. B. Inactivation of Salmonella in liquid egg albumen by antimicrobial bottle coatings infused with allyl isothiocyanate, nisin and zinc oxide nanoparticles. Appl. Microbiol.,2011, 110, 704–712. https://doi.org/10.1111/j.1365-2672.2011.04938.x.
CrossRef - Chen W., Jin T. Z., Gurtler J. B., Geveke D. J., Fan X. Inactivation of salmonella on whole cantaloupe by application of an antimicrobial coating containing chitosan and allyl isothiocyanate, J. Food Microbiol.,2012, 155, 165–170. https://doi.org/10.1016/j.ijfoodmicro.2012.02.001.
CrossRef - Maruthupandy M., Seo J. Allyl isothiocyanate encapsulated halloysite covered with polyacrylate as a potential antibacterial agent against food spoilage bacteria, Sci. Eng. C. Mater. Biol. Appl., 2019, 105, 110016. https://doi.org/10.1016/j.msec.2019.110016.
CrossRef - Banerjee A., Penna S., Variyar P. S. Allyl isothiocyanate enhances shelf life of minimally processed shredded cabbage, Food Chem., 2015, 183, 265–272. https://doi.org/10.1016/j.foodchem.2015.03.063.
CrossRef - Ugolini L., Righetti L., Carbone K., Paris R., Malaguti L., Di Francesco A., Micheli L., Paliotta M., Mari M., Lazzeri L. Postharvest application of Brassica meal-derived allyl-isothiocyanate to kiwifruit: effect on fruit quality, nutraceutical parameters and physiological response, Food Sci. Technol., 2017, 54, 751-760. https://doi.org/10.1007/s13197-017-2515-x.
CrossRef - Wu H., Xue N., Hou C. L., Feng J. T., Zhang X. Microcapsule preparation of allyl isothiocyanate and its application on mature green tomato preservation, Food Chem., 2015, 175, 344–349. https://doi.org/10.1016/j.foodchem.2014.11.149.
CrossRef - Ko J. A., Kim W. Y., Park H. J. Effects of microencapsulated Allyl isothiocyanate (AITC) on the extension of the shelf-life of Kimchi, J. Food Microbiol., 2012, 153, 92–98. https://doi.org/10.1016/j.ijfoodmicro.2011.10.021.
CrossRef - Amofa-Diatuo T., Anang D. M., Barba F. J., Tiwari B. K. Development of new apple beverages rich in isothiocyanates by using extracts obtained from ultrasound-treated cauliflower by-products: Evaluation of its physical properties and consumer acceptance, Food Compost. Anal., 2016, 61, 73-81. http://dx.doi.org/10.1016/j.jfca.2016.10.001.
CrossRef - Torrijos R., Nazareth T. M., Pérez J., Mañes J., Meca G. Development of a bioactive sauce based on oriental mustard flour with antifungal properties for pita bread shelf life improvement, Molecules,2019, 24, 1019. https://doi.org/10.3390/molecules24061019.
CrossRef - Lopes L., Meca G., Bocate K., Nazareth T., Bordin K., Luciano F. Development of food packaging system containing allyl isothiocyanate against Penicillium nordicum in chilled pizza: Preliminary study: LOPES etal., Food Process Preserv., 2017, 42, 13436. https://doi.org/10.1111/jfpp.13436.
CrossRef - Wu H., Zhang X., Zhang G., Zeng S. Y., Lin K. C. Antifungal vapour‐phase activity of a combination of allyl isothiocyanate and ethyl isothiocyanate against Botrytiscinerea and Penicilliumexpansum infection on apples, Phytopathol., 2011, 159, 450 – 455. https://doi.org/10.1111/j.1439-0434.2011.01792.x.
CrossRef - Recio R., Vengut-Climent E., Borrego L. G., Khiar N., Fernández I. Biologically active isothiocyanates: protecting plants and healing humans, Nat. Prod. Chem., 2017, 53, 167-242. https://doi.org/10.1016/B978-0-444-63930-1.00006-5.
CrossRef - Nadarajah D., Han J. H., Holley R. A. Inactivation of Escherichia coli O157:H7 in packaged ground beef by allyl isothiocyanate, Int. J. Food Microbiol.,2005, 99, 269–279. https://doi.org/10.1016/j.ijfoodmicro.2004.08.019.
CrossRef - Lin C. M., Preston J. F., Wei C. I. Antibacterial mechanism of allyl isothiocyanate, Food Prot., 2000, 63, 727–734. https://doi.org/10.4315/0362-028x-63.6.727.
CrossRef - Abbes C., Mansouri A., Landoulsi A. Synergistic effect of the lactoperoxidase system and cinnamon essential oil on total flora and salmonella growth inhibition in raw milk, Food Qual., 2018, 2018, 1-6. https://doi.org/10.1155/2018/8547954.63.6.727.
CrossRef - Al-Baarri A., Damayanti N., Legowo A., Tekiner İ., Hayakawa S. Enhanced antibacterial activity of lactoperoxidase–thiocyanate–hydrogen peroxide system in reduced-lactose milk whey, J. Food Sci., 2019, 2019, 1-6. https://doi.org/10.1155/2019/8013402.
CrossRef - Florence F., Grandison A., Lewis M. Factors affecting lactoperoxidase activity, J. Dairy Technol., 2005, 58, 233-236. https://doi.org/10.1111/j.1471-0307.2005.00227.x.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






