
Introduction
The intestine is a habitat for diverse micro-organisms that regulate intestinal homeostasis and immunological functions.1,2 The gut microbiota and intestinal immunity are inextricably connected, with a healthy microbiome promoting immune system development and function3,4 by the production of microbial metabolites such as short-chain fatty acids (SCFAs).5–12 Prebiotics are defined as “A non-digestible food ingredient that beneficially affects the host by selectively stimulating the growth and activity of bacteria in the colon and thus improves host health”.13–20
The research on the synergistic effects of okara-based prebiotics and L. plantarum probiotics highlights a promising yet underexplored area in gut health. Okara, or soybean residue, is a by-product of the tofu and soymilk manufacturing process. Despite its high quantity of nutritional fiber, protein, and beneficial components, it is often overlooked and thrown as waste. It is high in dietary fiber, specifically soluble and insoluble fibers and oligosaccharides, as well as proteins and other bioactive components such as isoflavones and antioxidants. The oligosaccharides in the okara serve as substrates for healthy gut flora, promoting their growth and activity. It holds immense potential as a prebiotic.21–24 These assist in protecting the gut lining from oxidative damage and inflammation. Okara additionally serves by lowering inflammation in the stomach and controlling immunological response, and it confers health advantages to hosts.25–32 Researchers reported the prebiotic effect of okara by carrying out in-vitro studies using L.acidophilus and Bifidobacterium bifidum33,34, and it was noted that okara provided a surface for the adhesion of cells, which ultimately enabled the uptake of the substrate and cell growth. Furthermore, insufficient research has been discovered regarding the precise mechanisms by which okara-derived prebiotics and L. plantarum interact, particularly their ability to improve probiotic viability, modify the composition of the gut microbiota, and improve overall gastrointestinal health. The current study addresses this research gap by integrating the probiotic efficacy of L. plantarum with the prebiotic potential of okara.
L.plantarum is a potential probiotic lactic acid bacterium commonly found in various fermented foods and the human gastrointestinal tract.35–37 According to research, L. plantarum strains have exceptional resistance to bile salts and hydrochloric acid (pH 2.0), with survival rates varying from 0.003 to 10%, in contrast to other strains such as L. sakei and L. paracasei.38 In vivo investigations have also demonstrated that L. plantarum has a high survival rate subsequent to human consumption. For instance, after a single injection of 108 cells, L. plantarum NCIMB 8826 had a 77% survival rate in the human ileum, much surpassing other strains like Lactococcus lactis and Lactococcus fermentum.39 Furthermore, L. plantarum can survive in the human gut for up to 11 days adhering to consumption, with colonization rates varying by individual.40 L. plantarum’s ability to attach to human cells, particularly via mannose-inhibited binding, is crucial for its survival and probiotic function. It may also protect the host against pathogenic infections.41,42
It possesses various probiotic qualities important for gut health, including resistance to acidic conditions in the stomach and bile salts in the intestine, improving gut survival and colonization. It also produces anti-inflammatory chemicals and modulates immunological responses, which reduce intestinal inflammation. The research aims to develop a synergistic synbiotic nutraceutical formulation containing L. plantarum as the probiotic and okara as a prebiotic high in dietary fiber. This composition attempts to increase the viability and stability of beneficial microbes, eventually promoting gut health by inhibiting the development of pathogenic bacteria. To accomplish this, the study focuses on improving the encapsulation of L. plantarum within okara, assuring the probiotic’s viability and efficacy. The formulation was further developed by evaluating the physicochemical parameters of freeze-dried powder formulation.
Furthermore, the stability of L. plantarum was investigated demonstrating the formulation’s viability and efficacy in maintaining a healthy gut microbiome.
Materials and Methods
Materials
L. plantarum was isolated from cabbage, a vegetable source. One gram of cabbage was suspended in saline, vigorously mixed, and centrifuged at 300 g to settle the particle matter. The supernatant was serially diluted tenfold with saline, and 100 μL of the 10-7 dilution was plated on MRS agar. The plates were then incubated at 30°C in microaerophilic conditions for 24-48 hours.43 Mprex Healthcare Pvt. Ltd. Waked, Pune, provided gift samples of okara (Soyabean waste). Lactobacillus de Man Rogosa and Sharpe (MRS) agar, MRS broth, McConkey agar, McConkey broth, fructooligosaccharide (FOS), sorbitol, mannitol, and bovine serum albumin (BSA) was acquired from HiMedia, Mumbai, India. Maltodextrin was sourced from Lobachemie, Mumbai, India. All reagents utilized in this investigation were high purity and fulfilled the analytical standards of the experimental techniques.
Phenotypic Characterization of L. plantarum
Characterization of L. plantarum
The L. plantarum was cultured on an MRS agar plate using the streak plate technique and incubated at 37℃ for 48 hr. The colony morphology, shape, colour, elevation, margin, and other notable features were examined. Gram staining was performed on a clean, grease-free slide to study the staining morphology. A smear of suspension was created, which was then heat-fixed onto the slide. The initial staining was performed with a water-soluble crystal violet dye, grams of iodine as a mordant, 95% ethyl alcohol as a decolourizing agent, and safranin as a counterstain.44,45 After drying, the smear was examined using an oil immersion objective.
Catalase Test and Oxidase Test
One small amount of well-isolated L. plantarum colony was mounted on a microscope slide. One drop of 2% H2O2 was added to the bacteria immediately and covered with a cover slip. The formation of effervescence Was observed after the addition of H2O2. Furthermore, the oxidase test was performed to determine whether the isolates could utilize cytochrome C oxidase, an enzyme in the electron transport chain.46 An isolated bacterial colony was transferred to a sterile Kovacs oxidase reagent (tetramethyl-p-phenylene-diamine dihydrochloride) oxidase test disc.47 The findings were reported based on colour changes; the formation of a deep purple/blue colour indicates oxidase production, while no change in colour indicates negative results.
TSI (Triple Sugar Iron) Test
The TSI medium contained three sugars (lactose, sucrose, and glucose) and iron, which are utilized to confirm the bacteria’s capacity to utilize these sugars. TSI agar slants were prepared and inoculated with L. plantarum by stabbing through the butt of the medium, then streaking the slants. The inoculated slant was kept at 37 °C for 18-24 hours. Following incubation, sugar fermentation, gas production, and hydrogen sulphide production were all examined with color change and gas production.46,48
Identification using the Vitek 2C system
The colonies of lactobacilli from MRS agar plates were identified using the Vitek 2C system (VITEK 2 Systems Version 9.2) following a preliminary screening. The ANC database includes a collection of 63 different types of anaerobic bacteria, including Lactobacillus.49,50 The individual lactobacilli colonies were selected from MRS agar plates and suspended in normal saline, exhibiting turbidity ranging from 2.7 to 3.3 McFarland units. The inoculums were introduced into an ANC (Anaerobic and Corynebacteria identification) card in the Vitek 2C system and incubated for approximately 6 hours. The identification process is initiated by examining microbial growth patterns and biochemical responses to determine the species and provide interpretations for final identifications.50
Characterization of Okara
The pharmacognostic profiles of okara were evaluated, including total ash value, water-insoluble ash value, and acid-soluble ash value. The total ash value was determined by utilizing pre-weighed 2 g of okara incinerated in the furnace until constant weight was obtained. The water-insoluble ash value was examined using distilled water, and the acid-soluble ash value was determined using 10% HCl.51 The phytochemical screening procedures of okara comprise the molish test for carbohydrate content, the biuret test for protein content, and the bromine water test for tannin content.
Bradford assay
The Bradford assay determined the protein content of the okara in comparison to BSA, which was used as the reference standard to plot a calibration curve using known concentrations.52 The okara and BSA solutions were prepared, resulting in stock solutions with a concentration of 2000 µg/mL in distilled water. To achieve concentrations of 100 µg/mL, 200 µg/mL, 400 µg/mL, 600 µg/mL, 800 µg/mL, 1000 µg/mL, and 1200 µg/mL, samples were collected from test solutions and diluted with distilled water to a final volume of 10 mL. Bradford dye solution (Coomassie brilliant blue G-250) (5 mL) was added to each diluted sample, and the solutions were incubated for 5 to 10 min. The absorbance was measured at 595 nm after incubation.53
Prebiotic efficacy of okara
The growth kinetics of L. plantarum were evaluated in reconstituted MRS broth supplemented with okara and FOS and containing tryptone, pectin, meat extract, potassium phosphate dibasic, sodium acetate, tri-ammonium citrate, and magnesium sulphate. L. plantarum cultured in MRS broth was utilized as the blank sample. To cultivate L. plantarum, 2% (v/v) inoculum was added to 50 mL of sterile, reconstituted MRS broth under aseptic conditions. The flasks were subsequently placed in an orbital shaking incubator (Remi electro tech, CIS 18, India) and incubated for 48 hours at 37± 2 °C. Samples were collected after 0, 2, 4, 6, 24, and 48 hours, and the optical density (OD) at 600 nm was recorded. The growth rate (μ) was calculated during the exponential phase of growth using the following formula,
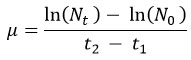
Where,
μ = growth rate (h⁻¹)
Nt = Bacterial population at time t2
N0 = Bacterial population at time t1
t2 – t1 = time interval between the two measurements
The doubling time (g) was calculated from the growth rate using the following formula,
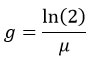
A cooling centrifuge (Remi electro tech, C-24 plus ZFCU-07420, India) was used to centrifuge the culture at 1000 rpm to produce a cell-free supernatant for further analysis. The bacterial pellet that remained over after the centrifugation was utilized to evaluate the dry biomass.
Determination of dry biomass
To monitor the cellular development process, a cooling centrifuge (Remi electro tech, C-24 plus ZFCU-07420, India) was used to separate the developed bacterial cells pellet by centrifuging the sample at 1000 rpm for 30 minutes. The separated cell pellet was dried in a hot air oven at 40 °C (Remi Electro Tech, AI-7982, Mumbai, India) until it reached a consistent weight as dry biomass.
Determination of pH and Titratable Acidity
The pH of the cell-free supernatant was determined using a calibrated electronic digital pH meter (EQ-614A Equip-Tronic, Mumbai, India) at each time interval. This method indicates organic acid generation and is essential for determining the Lactobacillus probiotic and fermentative characteristics.
The % Titratable acidity of L. plantarum was measured by titrating the bacterial cell-free supernatant with 0.1 M NaOH and using phenolphthalein as an indicator. The titratable acidity was calculated as follows,
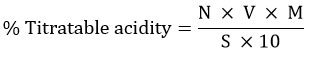
Where, N is the Normality of NaOH, V is the Volume of NaOH used for titration (mL), and M is the Molecular weight of the predominant acid in the sample divided by the number of hydrogen ions in the acid molecules that are titrated, S is the Sample size in mL or g.
Determination of anti-microbial activity
The antibacterial activity of cell-free supernatant from L. plantarum culture was assessed against E. coli using the agar well diffusion technique using MacConkey agar. The overnight cultures (0.1 mL ) of E. coli were evenly distributed on agar. The wells were cut to a diameter of 6mm, and each well received 0.1 mL of supernatant. After allowing the sample to diffuse for 4 hours in the refrigerator, plates were incubated at 37 ℃ for 24 hours under aerobic conditions. The zone of inhibition was calculated.
Formulation of freeze-dried powder
The formulation was performed on a laboratory-size lyophilizer (Martin Christ, Alpha 2-4LSC, An der Unteren Sose, Germany). The formulation was optimized for the carrier type (mannitol, sorbitol, and maltodextrin) and carrier-to-prebiotic ratio (0.5:1, 1:1, 2:1), as shown in the table. Okara was used as a prebiotic, with mannitol, sorbitol, and maltodextrin serving as cryoprotectants for lyophilization. L. plantarum was utilized as a probiotic at 1.2×109 CFU/mL. The samples were lyophilized for 30 minutes for freezing, warming for 10 minutes, primary drying for 4 hours (vacuum 0.0050 m pressure), and final drying for 3 hours (vacuum 0.1000 m bar). The process was continued for two days to eliminate any residual moisture.
Formulation of synbiotic freeze-dried powder
Nine batches were evaluated for powder yield and micromeritic parameters. These investigations were performed to select the batch with the highest yield and optimal micromeritic properties. Each batch underwent evaluation under the same conditions, and the results were compared to determine the optimal powder yield and appropriate micromeritic characteristics.
Powder yield and micromeritic properties
The yield of freeze-dried powder was determined by dividing the weight of freeze-dried powder by the theoretical weight of solids before freeze-drying.
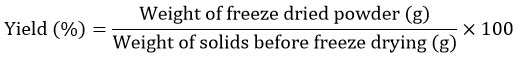
The micromeritic properties of the freeze-dried powder were evaluated in compliance with the USP 2018 standard methodology, including the Bulk density, Taped density, Carr’s index, Hausner’s ratio, and Angle of repose.
Particle size
Optical microscopy
Particle size was measured in freeze-dried formulations using optical microscopy and a calibrated eyepiece micrometer. The freeze-dried powder on the slide and the size of 100 particles were measured under the microscope using the eyepiece scale, which was divided into 100 minor divisions. Each division is equivalent to 10 meters. The arithmetic and geometric mean diameters of the particles were calculated.
Laser Diffraction
Laser diffraction (Malvern Meta sizer) was used to measure the particle size of the optimized freeze-dried powder batch. A cumulative undersize was created, allowing the identification of granule sizes corresponding to the 10th, 50th, 90th, and 100th percentiles (D10, D50, D90, and D100). The calculation was expressed in terms of arithmetic mean diameter.
Evaluation of reconstituted powder
Viability enumeration
The formulation’s viability was tested before and after freeze-drying by spreading it on MRS agar plates. The results are expressed in CFU/mL using mean and standard deviation. The viability of freeze-dried powder was determined by performing ten-fold serial dilutions of reconstituted powders on MRS agar plates. The reconstituted powder was serially diluted with MRS broth before plating. The MRS plates were incubated at 37 °C for 48 hours, and the bacterial colonies were counted.
In-vitro efficacy using the co-culture technique
Minimal Essential Medium (MEM) was prepared, which included yeast extract, tween 80, tryptone, meat extract, potassium phosphate dibasic, sodium acetate, triammonium citrate, and magnesium sulphate. The media was then inoculated with 0.5 MacFarland standards of E. coli. Several flasks were prepared: flask 1 contained the control E. coli, flask 2 contained the formulation and E. coli, flask 3 contained L. plantarum, and flask 4 contained L. plantarum with formulation and E. coli. The flasks were incubated for 48 hours. At 0, 6, 24, and 48 hours of incubation, the sample was withdrawn from each flask and added to the sterilized broth to make serial dilutions (10-8, 10-10, 10-12). They were subjected to a 24-hour incubation period at 37 ℃. MRS agar was used for the formulation containing L. plantarum, and MacConkey agar for E. coli. The colony count (CFU/mL) was calculated using the formula as follows,
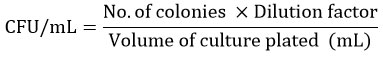
Statistical analysis
Data from each experiment was reported as mean ± standard error mean (SEM). The statistical significance was determined using Graph pad prism software (version 6.0, USA) to assess the statistical significance of the data by one-way analysis of variance (ANOVA) followed by Tukey’s multiple comparison test. The control group was compared with the other groups. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, ns: non-Significant.
Results
Standardization of L. plantarum
After incubation, the L. plantarum colonies demonstrated a creamy white opaque colour. The appeared colonies were small, round, and exhibited a raised elevation with an entire margin.
Catalase Test and Oxidase Test
The catalase test was negative, as no bubbles were observed. L.plantarum restricts the breakdown of H2O2 and further leads to the accumulation of H2O2 within the bacterial cell, which can limit the proliferation of pathogenic micro-organisms. In the case of the oxidase test, no change was observed on the oxidase disc within 15-30 seconds after adding an isolated bacterial colony, indicating negative oxidase test results.
Triple Sugar Iron test
The L. plantarum TSI test results showed an alkaline butt (red) and an acid slant (yellow), with no gas or H2S production. L.plantarum can aerobically ferment lactose and sucrose to generate acid, as evidenced by the yellow slant.
VITEK analysis
The biochemical makeup of L. plantarum was examined using the VITEK 2C identification technology.54 L. plantarum provided several satisfactory findings, which are essential for species identification. Aminopeptidases, including leucine, proline, and tyrosine, verified L. plantarum’s distinctive enzymatic profile (Table 1). The outcomes additionally displayed beta-glucoside activity and alpha-mannosidase activity.
Table 1: The biochemical reactions for the diagnostic features of L.plantarum using Vitek 2C identification
| Biochemical Test | Reaction* |
| dGAL (D-Galactose) | – |
| LeuA (Leucine Aminopeptidase) | + |
| ELLM (L-Ellman) | – |
| PheA (Phenylalanine Aminopeptidase) | + |
| ProA (Proline Aminopeptidase) | + |
| PyrA (Pyrrolidonyl Aminopeptidase) | – |
| dCEL (D-Cellobiose) | + |
| TyrA (Tyrosine Aminopeptidase) | + |
| APPA (L-Alanine Aminopeptidase) | – |
| dGLU (D-Glucose) | + |
| dMNE (D-Mannitol) | + |
| dMAL (D-Maltose) | + |
| SAC (Sucrose) | + |
| ARB (L-Arabitol) | + |
| NAG (N-Acetyl-Glucosamine) | + |
| BGLUi (Beta-Glucosidase) | + |
| URE (Urease) | – |
| BGURi (Beta-Glucuronidase) | – |
| BGALi (Beta-Galactosidase) | – |
| AARA (Alpha-Arabinosidase) | – |
| AGALi (Alpha-Galactosidase) | – |
| BMAN (Beta-Mannosidase) | – |
| ARG (Arginine Dihydrolase) | – |
| PVATE (Phosphatidylinositol Phospholipase C) | – |
| MTE (Alpha-Mannosidase) | + |
| ESC (Esculin Hydrolysis) | + |
| BdFUC (Beta-D-Fucosidase) | – |
| BNAGi (Beta-N-Acetylglucosaminidase) | – |
| AMANi (Alpha-Mannosidase) | – |
| AIFUC (Alpha-L-Fucosidase) | – |
| PHOS (Phosphatase) | – |
| IARA (Inositol) | – |
| dRIB2 (D-Ribose) | + |
| OPS (Ornithine Decarboxylase) | – |
| AARAF (Alpha-L-Arabinofuranosidase) | – |
| dXYL (D-Xylose) | – |
| GRAM (Gram Stain) | + |
| MORPH (Morphology) | – |
| AERO (Aerobic Growth) | ? |
* (+) Positive enzyme activity / Positive fermentation, (-) No activity / No fermentation, (?) Uncertain
Characterization of okara
Pharmacognostic evaluations
The pharmacognostic characteristics of okara revealed a total ash value of 4 ± 0.02%, an acid-insoluble value of 1 ± 0.01%, and a water-soluble ash value of 3.75% which was within the acceptable range and is not more than the standard value, i.e., 5%.
Bradford assay of okara
The present investigation utilized the Bradford test to assess okara’s protein content quantitatively. BSA was utilized as a standard. The BSA standard curve was obtained with R2 = 0.9948 and the equation (y = 0.0006x + 1.1384). The range of protein concentrations in okara was 52.25 µg/mL to 1042.25 µg/mL, where higher absorbance values corresponded to more significant protein concentrations.
Prebiotic efficacy of okara
Growth kinetics of okara
The growth kinetics were investigated using resubstituted MRS broth with okara at various concentrations (1%, 2%, 3%, and 4%), L. plantarum as a probiotic, and FOS as a comparison sugar source. The okara exhibited robust growth at 2% of okara concentration with the specific growth rate (μ) at 0.064 at 2 h, and it increased up to 1.73 at 48 h, demonstrating significant and sustained growth (Table 2).
Table 2: Specific Growth Rate of L. plantarum in response to varying concentrations of okara as a prebiotic over different time intervals.
| Time (hr) | 1% | 2% | 3% | 4% | FOS | Control |
| μ2 | 0.044 | 0.064 | 0.096 | 0.075 | 0.068 | 0.0175 |
| μ4 | 0.064 | 0.100 | 0.148 | 0.1625 | 0.149 | 0.0295 |
| μ6 | 0.084 | 0.323 | 0.177 | 0.211 | 0.517 | 0.103 |
| μ24 | 0.546 | 0.662 | 0.866 | 0.755 | 1.225 | 0.478 |
| μ48 | 0.918 | 1.737 | 1.209 | 1.016 | 1.896 | 1.245 |
| μavg | 0.3312 | 0.5772 | 0.4992 | 0.4439 | 0.771 | 0.3746 |
pH and % titratable acidity
l. plantarum can produce organic acids by fermentation of sugars, and a majority of lactic acid is produced. Initially, it was neutral to partially alkaline. The amino acid composition of okara can contribute to an increase in pH. Among all the okara concentrations, 2% was the most effective at promoting bacterial growth and lactic acid production. Although 3% okara produced a significant pH reduction, 1% and 4% concentrations had less influence on pH (Figure 1b).
The % titratable acidity for L. plantarum gradually increased with time and various concentrations of okara. At 0 h, acidity levels were relatively low across all concentrations. Acidity gradually increased during incubation. By 48 hours, the bacteria had optimum acidity levels of 2% (0.94) and 3% (0.87), respectively, with FOS having the highest acidity at 1.10 and the control at 0.92 representing L.plantarum’s highest metabolic activity and acid production (Figure 1c).
Dry biomass
Dry biomass concentrations (g/L) of L. plantarum were determined in various concentrations of okara, fructooligosaccharides (FOS), and control conditions periodically. All concentrations had no dry biomass at 0, 2, and 4 hours. As described in Figure 1d, at 6 hours, biomass started to accumulate. Notably, 2% okara and FOS produced greater biomass than the other concentrations, with values of 0.34 and 0.58 g/L, respectively. In contrast, the remaining okara concentrations (1%, 3%, and 4%) and the control had lower biomass concentrations throughout the observation period, indicating slower biomass accumulation.
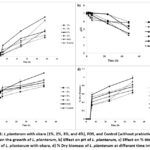 |
Figure 1: L.plantarum with okara (1%, 2%, 3%, and 4%), FOS, and Control (without prebiotic) for a) Effect on the growth of L. plantarum, b) Effect on pH of L. plantarum, |
Anti-microbial activity
The zone of inhibition (ZOI) increased significantly for all lactobacillus samples between 6 and 48 h, indicating a time-dependent effect (Figure 2a). After 6 h, the 7.1 ± 0.3 mm ZOI showed significant inhibition. The ZOI rose after 24 h by 9.2 ± 0.3 mm, indicating greater susceptibility to E. coli. The ZOI significantly increased after 48 h up to 16.9 ± 0.2 mm, demonstrating that E. coli is responsive to Lactobacillus’ anti-microbial effect.
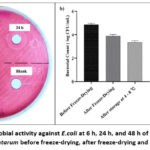 |
Figure 2: a) Anti-microbial activity against E.coli at 6 h, 24 h, and 48 h of exposure, b) Viability enumeration of L. plantarum before freeze-drying, after freeze-drying and after storage at 4-8 ℃. |
Evaluation of Freeze-dried Powder
A novel formulation of L. plantarum and okara has been developed as a freeze-dried powder. With the incorporation of sugars like mannitol, sorbitol, and maltodextrin. 9 batches were developed by adding okara, mannitol, sorbitol, and maltodextrin with probiotics with different ratios. The 3 best batches with the highest yields were selected for further evaluation from the nine batches.
The three scrutinized batches (B1, B2, and B4) were examined for yield, flow properties, and particle size metrics to determine the most optimal batch after lyophilization of freeze-dried powder. Batch B1 yielded 85.2% with a 2:1 okara-mannitol ratio, Batch B2 yielded 84.6% with the same ratio, and Batch B4 yielded the highest yield at 86.6%. While Batch B4 had the highest yield due to the okara: sorbitol ratio (2:1), Batch B2 was distinguished by its good flow and particle size characteristics. Batch B2 exhibits a less significant drying loss (2.1%) than Batch B4 (8.6%), indicating higher stability and reduced moisture retention. This is essential to maintaining powder quality throughout the process. Batch B2 exhibited less agglomeration than Batch B1 (34.9°) and Batch B4 (30.9°), with the lowest angle of repose (26.5°) indicating better flow.
B1 exhibited larger particle sizes than Batch B2 (87.7 µm and 82.03 µm, respectively) (Table 3). The arithmetic mean for Batch B4 is 85.15 µm, and the geometric mean was 82.53 µm. Smaller particle sizes increase flowability and reduce agglomeration. Furthermore, laser diffraction (Malvern Mastersizer 2000) was used to determine the particle size of batch B2. A cumulative undersize curve was created, enabling the identification of granule sizes at the 10th, 50th, 90th, and 100th percentiles (D10, D50, D90, and D100). The estimation covered a range of 4.537 µm to 434.390 µm.
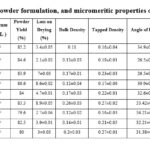 |
Table 3: Composition of the powder formulation, and micromeritic properties of the powder formulation. |
Evaluation of reconstituted freeze-dried powder
Viability Enumeration
The objective of the study was to determine the vitality of bacterial cells during various stages of preservation, particularly before and after freeze-drying, as well as after storage at 4 – 8 °C. Before freeze-drying, the initial bacterial count was 4.06 ± 0.023 log CFU/mL (or 4.66 ± 0.023 log CFU/g). The bacterial count decreased to 3.78 ± 0.045 log CFU/mL (or 4.40 ± 0.045 log CFU/g) after the freeze-drying procedure, suggesting a slight reduction in viability. A bacterial count of 3.54 ± 0.015 log CFU/mL (or 4.16 ± 0.015 log CFU/g) was obtained after additional storage at 4 – 8 °C for 7 days, indicating a progressive decrease in viability over the storage time (Figure 2b).
In-vitro efficacy by co-culture technique
A mixed culture technique was used to study the effect of synbiotic freeze-dried powder formulation on E. coli. The formulation with probiotic and prebiotic was incubated alone in MRS media to observe the growth of L. plantarum and in the presence of E. coli culture to study the mixed culture effect. When the formulation was incubated in media without any pathogen, there was a sustained increase in L. plantarum at 24 h (8.26 ± 0.12 log CFU/mL) and 48 h (9.68 ± 0.19 log CFU/mL). The bacterial colony count at 0 h was found to be 5.96 ± 0.06 CFU/mL for group 1 (control of E. coli), which increased to 7.22 ± 0.09 CFU/mL at 24 h and 8.20 ± 0.05 CFU/mL at 24 h. Group 2 (Formulation + E. coli) showed 5.37 ± 0.08 CFU/mL initially, while at 24 h, the colony count increased to 6.31 ± 0.05 CFU/mL and 7.25 ± 0.09 CFU/mL at 48 h. For Group 3 (Control of L. plantarum), the colony count observed was 6.06 ± 0.13 CFU/mL at 0 h, gradually increasing to 7.76 ± 0.27 at 24 h and 8.64 ± 0.21 at 48 h. In group 4, containing only formulation, the colony count was observed as 5.98±0.02 CFU/mL at 0 h, 8.26 ± 0.12 CFU/mL at 24 h and 9.68 ± 0.19 CFU/mL at 48 h (Figure 3b).
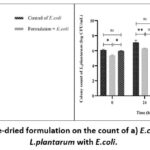 |
Figure 3: Effect of freeze-dried formulation on the count of a) E.coli and b) Co-culture of L.plantarum with E.coli. |
Data were expressed as mean ± SEM, n=3 and Tukey’s Multiple comparisons analysis by two-way analysis of variance (ANOVA). The control group is compared with the other groups.
*p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, ns: non-significant.
Discussion
The staining observed thick peptidoglycan layers.45 and the presence of gram-positive bacilli with the characteristic L. plantarum staining appearance, which showed purple-coloured, rod-shaped bacteria with single rods, pairs, and short chains with apparent length variations.
The catalase enzyme neutralized the bactericidal effects of hydrogen peroxide (H2O2). It promotes the breakdown of H2O2 into water and oxygen and also inhibits the antibacterial properties of hydrogen peroxide, and its quantity in bacteria has been associated with pathogenicity.55–57 In case of oxidase test, L. plantarum lacks cytochrome c oxidase. This absence enabled the L. plantarum to survive in anaerobic or low-oxygen conditions with the ability to produce acidic conditions that inhibit the proliferation of pathogenic micro-organisms. The crimson buttocks indicated that the L. plantarum fermented glucose anaerobically in TSI. The absence of blackening indicated the absence of H2S generation, whereas the absence of bubbles confirms the absence of gas production.
In VITEK analysis the bacterium can be recognized easily because of its esculin hydrolysis, absence of urease production, and negative reactivity to other enzyme activities such as beta-glucuronidase, beta-galactosidase, and arginine dihydrolase. These tests are biochemical indicators that distinguish the L. plantarum from the other lactobacillus species. The L. plantarum profile showed various types of carbohydrates, including D-cellobiose, D-glucose, D-mannitol, D-maltose, sucrose, L-arabitol, N-acetyl-glucosamine, and D-ribose. These demonstrate the fermentation ability of L. plantarum. All the biochemical characteristics confirmed the presence of L. plantarum precisely.
The pharmacognostic tests of okara yielded positive confirmatory tests such as the molish test for carbohydrates, the biuret test for proteins, and the bromine water test for tannins. In the case of the growth kinetics of okara, the doubling time (g) for 2% was 0.39 h at 48 h. Similarly, FOS exhibited a consistent, substantial proliferation with a specific growth rate of 1.89 at 48 h and doubling time at 0.36 h, confirming its ability to promote bacterial growth. In contrast, the control exhibited gradual development with an initial specific growth rate of 0.017 at 2 h and increased to 1.24 after 48 h (Figure 1a). The doubling time for control was 0.55 h at 48 h, indicating that the L. plantarum growth was substantially less supported without adding okara. Other okara concentrations showed a relatively small increase, with an initial specific growth rate of 0.044 at 2 h and 0.91 at 48 h, corresponding doubling time of 0.75. The 3% okara concentration proliferated faster than 1% but less effectively than 2%. Additionally, 4% okara showed noteworthy growth, with an initial specific growth rate of 1.016 at 48 h, with a doubling time of 0.68 h at 48 h. A significant increase was provided by 2 % okara and FOS, which indicated more excellent specific growth rates and shorter doubling time, indicating its efficacy as a prebiotic.
The pH declined substantially from 7 to 4.24 after 48 h, indicating significant fermentation activity and optimum conditions for L. plantarum. Correspondingly, FOS presented a considerable pH decrease, representing that the bacteria produced acid rapidly. In contrast, the control group (without prebiotics) had the minimum variation in pH, initially at 6.9 and declining gradually to 5.8 at 48 h. This emphasizes the role of okara as a prebiotic and FOS in L. plantarum’s growth and metabolic activity.
Although Batch B4 has the shortest arithmetic mean particle size, Batch B2’s geometric mean particle size is also reasonably small, indicating strong flow properties without compromising stability. While Batch B1 had the highest output, its flow properties are less favourable, with a Carr’s Index of 52.94 and a higher angle of repose, making it more prone to handling difficulties. Despite having a higher yield and better particle size distribution, Batch B4 had a higher drying loss and less favourable flow properties than Batch B2. Batch B2 was selected for its balanced combination of low drying loss, good flow properties, and relatively small particle size.
Viability Enumeration findings implied that although bacterial viability is affected by freeze-drying and subsequent cold storage, the effects were minor. Consequently, freeze-drying and storage at 4 – 8 °C for 7 days can be considered an effective method to preserve bacteria alive, even if there will always be some bacterial count reduction.
In-vitro efficacy by co-culture technique showed steady growth at 24 h and 48 h control with E. coli (Figure 3a). When the formulation was incubated in E. coli culture, there was an increase in L. plantarum growth and a decrease in the growth of E. coli. This inferred that E. coli does not inhibit L. plantarum, whereas prebiotics support the development of L. plantarum.
Conclusion
The current research successfully developed a synbiotic nutraceutical formulation containing L. plantarum as a probiotic and okara as a prebiotic, using freeze-drying process. Batch B2 emerged as the most preferable among the nine selected batches because of its optimal properties. Okara demonstrated the potential prebiotic efficacy through growth kinetics of L. plantarum, by significantly enhancing its growth rate and biomass accumulation. The viability studies confirmed that the freeze-drying process effectively upheld the viability of bacterial cells, with minimal reduction in bacterial count after storage. The formulation’s in-vitro efficacy showed substantial growth of L. plantarum and inhibition of E. coli, indicating the prebiotic’s role in promoting probiotic growth and suppressing pathogens. In conclusion, the study highlights the successful development and optimization of a synbiotic nutraceutical formulation that combines the benefits of probiotics and prebiotics, offering significant potential for enhancing gut health and contributing to improved nutrition.
Future studies will involve in-vivo animal trials to assess the efficiency of soybean-based synbiotic powder containing L. plantarum in enhancing gut health and altering gut microbiota. Long-term investigations could determine its preventative and therapeutic efficacy against gastrointestinal diseases. Furthermore, clinical trials could test its application in humans, ensuring scalability and application effectiveness.
Acknowledgement
We are thankful to Mprex Healthcare, Wakad, Pune for providing gift samples of the prebiotic okara. Sincere thanks to the Department of Science and Technology (DST) for the facility of the instrument established through the Fund of Improvement of S&T Infrastructure (FIST) Program at the institute from the Government of India, Department of Science and Technology, R and D Infrastructure Division, which is employed in this study.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability
All the data is available with the author and shall be provided upon request
Ethical Statement
This research did not involve human participants, animal subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Clinical Trial Registration
This research does not involve any clinical trials.
Permission to reproduce material from source
Not applicable.
Author Contributions
- Roshani Rajendra Pagar: Writing – original draft, Visualization, Software, Methodology, Investigation, Formal analysis, Data curation.
- Sanjeevani Deshkar: Conceptualization, Design of study, Revising and approval of manuscript, Validation, Supervision, and Data curation.
- Rewati Choudhary: Methodology, Investigation, Formal analysis.
- Dheeraj Nagore: Project management, Materials and Resources.
- Nandkishor Bhikane: Project management, Supervision.
References
- Milani C, Duranti S, Bottacini F, et al. The First Microbial Colonizers of the Human Gut: Composition, Activities, and Health Implications of the Infant Gut Microbiota. Microbiol Mol Biol Rev. 2017;81(4):e00036-17. doi:10.1128/MMBR.00036-17
CrossRef - Takiishi T, Fenero CIM, Câmara NOS. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers. 2017;5(4):e1373208. doi:10.1080/21688370.2017.1373208
CrossRef - Usuda H, Okamoto T, Wada K. Leaky Gut: Effect of Dietary Fiber and Fats on Microbiome and Intestinal Barrier. Int J Mol Sci. 2021;22(14):7613. doi:10.3390/ijms22147613
CrossRef - Leonardi I, Gao IH, Lin WY, et al. Mucosal fungi promote gut barrier function and social behavior via Type 17 immunity. Cell. 2022;185(5):831-846.e14. doi:10.1016/j.cell.2022.01.017
CrossRef - Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr. 1995;125(6):1401-1412. doi:10.1093/jn/125.6.1401
CrossRef - Thomas CM, Versalovic J. Probiotics-host communication: Modulation of signaling pathways in the intestine. Gut Microbes. 2010;1(3):148-163. doi:10.4161/gmic.1.3.11712
CrossRef - Bron PA, van Baarlen P, Kleerebezem M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat Rev Microbiol. 2011;10(1):66-78. doi:10.1038/nrmicro2690
CrossRef - Collado MC, Meriluoto J, Salminen S. Role of commercial probiotic strains against human pathogen adhesion to intestinal mucus. Lett Appl Microbiol. 2007;45(4):454-460. doi:10.1111/j.1472-765X.2007.02212.x
CrossRef - O’Shea EF, Cotter PD, Stanton C, Ross RP, Hill C. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. International Journal of Food Microbiology. 2012;152(3):189-205. doi:10.1016/j.ijfoodmicro.2011.05.025
CrossRef - Spinler JK, Taweechotipatr M, Rognerud CL, Ou CN, Tumwasorn S, Versalovic J. Human-derived probiotic Lactobacillus reuteri demonstrate antimicrobial activities targeting diverse enteric bacterial pathogens. Anaerobe. 2008;14(3):166-171. doi:10.1016/j.anaerobe.2008.02.001
CrossRef - Lee BJ, Bak YT. Irritable Bowel Syndrome, Gut Microbiota and Probiotics. J Neurogastroenterol Motil. 2011;17(3):252-266. doi:10.5056/jnm.2011.17.3.252
CrossRef - Venegas DP, Fuente MKD la, Landskron G, et al. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Frontiers in Immunology. 2019;10. doi:10.3389/fimmu.2019.00277
CrossRef - Davani-Davari D, Negahdaripour M, Karimzadeh I, et al. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods. 2019;8(3):92. doi:10.3390/foods8030092
CrossRef - Pokusaeva K, Fitzgerald GF, van Sinderen D. Carbohydrate metabolism in Bifidobacteria. Genes Nutr. 2011;6(3):285-306. doi:10.1007/s12263-010-0206-6
CrossRef - Kuo SM. The Interplay Between Fiber and the Intestinal Microbiome in the Inflammatory Response. Advances in Nutrition. 2013;4(1):16-28. doi:10.3945/an.112.003046
CrossRef - Hotel A. Health and Nutritional Properties of Probiotics in Food Including Powder Milk with Live Lactic Acid Bacteria – Joint FAO/WHO Expert Consultation. 2001;2014.
- Panda S, Behera DrS, Qaku WX, et al. Quality enhancement of prickly pears (Opuntia sp.) juice through probiotic fermentation using Lactobacillus fermentum – ATCC 9338. LWT – Food Science and Technology. 2016;75. doi:10.1016/j.lwt.2016.09.026
CrossRef - KHAN RU. The applications of probiotics in poultry production. World’s Poultry Science Journal. 2013;69. doi:10.1017/S0043933913000627
CrossRef - Fuller R. Probiotics in man and animals. J Appl Bacteriol. 1989;66(5):365-378.
CrossRef - Schrezenmeir J, de Vrese M. Probiotics, prebiotics, and synbiotics–approaching a definition. Am J Clin Nutr. 2001;73(2 Suppl):361S-364S. doi:10.1093/ajcn/73.2.361s
CrossRef - Swallah MS, Fan H, Wang S, Yu H, Piao C. Prebiotic Impacts of Soybean Residue (Okara) on Eubiosis/Dysbiosis Condition of the Gut and the Possible Effects on Liver and Kidney Functions. Molecules. 2021;26(2):326. doi:10.3390/molecules26020326
CrossRef - Asghar A, Afzaal M, Saeed F, et al. Valorization and food applications of okara (soybean residue): A concurrent review. Food Science & Nutrition. 2023;11. doi:10.1002/fsn3.3363
CrossRef - Feng JY, Thakur K, Ni ZJ, et al. Effects of okara and vitamin B2 bioenrichment on the functional properties and in vitro digestion of fermented soy milk. Food Research International. 2021;145:110419. doi:10.1016/j.foodres.2021.110419
CrossRef - Simoes I, Mustapha A. Okara as a novel prebiotic ingredient in the manufacture of a probiotic yogurt. http://undergradresearch. missouri.edu/ forums-conferences/abstracts/abstract-detail.php?abstractid=. Published online 2004. Accessed October 13, 2024. https://mospace.umsystem.edu/xmlui/handle/10355/1968
- Lesa KN, Ahmad N, Mayangsari Y, et al. Health Benefits of Okara for the Management of Diabetes Mellitus. Journal of Food Quality. 2023;2023(1):5540118. doi:10.1155/2023/5540118
CrossRef - Nguyen LT, Nguyen TH, Nguyen LT, et al. Okara Improved Blood Glucose Level in Vietnamese with Type 2 Diabetes Mellitus. J Nutr Sci Vitaminol (Tokyo). 2019;65(1):60-65. doi:10.3177/jnsv.65.60
CrossRef - Lesa KN, Ahmad N, Mayangsari Y, Cahyanto MN, Saputra WD. Anti-Diabetic Effect of Okara Noodles on Streptozotocin-Nicotinamide Induced Diabetic Rats. Trends in Sciences. 2024;21(5):7428-7428. doi:10.48048/tis.2024.7428
CrossRef - Saad N, Delattre C, Urdaci M, Schmitter JM, Bressollier P. An overview of the last advances in probiotic and prebiotic field. LWT – Food Science and Technology. 2013;50(1):1-16. doi:10.1016/j.lwt.2012.05.014
CrossRef - Parnell JA, Raman M, Rioux KP, Reimer RA. The potential role of prebiotic fibre for treatment and management of non-alcoholic fatty liver disease and associated obesity and insulin resistance. Liver Int. 2012;32(5):701-711. doi:10.1111/j.1478-3231.2011.02730.x
CrossRef - Zhang J, Wang S, Wang J, et al. Insoluble Dietary Fiber from Soybean Residue (Okara) Exerts Anti-Obesity Effects by Promoting Hepatic Mitochondrial Fatty Acid Oxidation. Foods. 2023;12(10):2081. doi:10.3390/foods12102081
CrossRef - Lemes SF, Lima FM, de Almeida APC, et al. Nutritional recovery with okara diet prevented hypercholesterolemia, hepatic steatosis and glucose intolerance. Int J Food Sci Nutr. 2014;65(6):745-753. doi:10.3109/09637486.2014.898259
CrossRef - Kamal E, Kaddam LA, Alagib A, Saeed A. Dietary Fibers (Gum Arabic) Supplementation Modulates Hepatic and Renal Profile Among Rheumatoid Arthritis Patients, Phase II Trial. Front Nutr. 2021;8:552049. doi:10.3389/fnut.2021.552049
CrossRef - Espinosa I, Rupérez P. Indigestible fraction of okara from soybean: Composition, physicochemical properties and in vitro fermentability by pure cultures of Lactobacillus acidophilus and Bifidobacterium bifidum. European Food Research and Technology. 2009;228:685-693. doi:10.1007/s00217-008-0979-7
CrossRef - Vieira ADS, de Souza CB, Padilha M, et al. Impact of a fermented soy beverage supplemented with acerola by-product on the gut microbiota from lean and obese subjects using an in vitro model of the human colon. Appl Microbiol Biotechnol. 2021;105(9):3771-3785. doi:10.1007/s00253-021-11252-8
CrossRef - Arasu MV, Al-Dhabi NA, Ilavenil S, Choi KC, Srigopalram S. In vitro importance of probiotic Lactobacillus plantarum related to medical field. Saudi J Biol Sci. 2016;23(1):S6-S10. doi:10.1016/j.sjbs.2015.09.022
CrossRef - de Vries MC, Vaughan EE, Kleerebezem M, de Vos WM. Lactobacillus plantarum—survival, functional and potential probiotic properties in the human intestinal tract. International Dairy Journal. 2006;16(9):1018-1028. doi:10.1016/j.idairyj.2005.09.003
CrossRef - (PDF) Lactobacillus plantarum: An overview with emphasis in biochemical and healthy properties. In: ResearchGate. Accessed November 21, 2024. https://www.researchgate.net/publication/272415707_Lactobacillus_ plantarum_An_overview_ with_emphasis _in_biochemical_and_healthy_properties
- Haller D, Colbus H, Gänzle MG, Scherenbacher P, Bode C, Hammes WP. Metabolic and Functional Properties of Lactic Acid Bacteria in the Gastro-intestinal Ecosystem: A comparative in vitro Studybetween Bacteria of Intestinal and Fermented Food Origin. Systematic and Applied Microbiology. 2001;24(2):218-226. doi:10.1078/0723-2020-00023
CrossRef - Vesa T, Pochart P, Marteau P. Pharmacokinetics of Lactobacillus plantarum NCIMB 8826, Lactobacillus fermentum KLD, and Lactococcus lactis MG 1363 in the human gastrointestinal tract. Aliment Pharmacol Ther. 2000;14(6):823-828. doi:10.1046/j.1365-2036.2000.00763.x
CrossRef - Johansson ML, Molin G, Jeppsson B, Nobaek S, Ahrné S, Bengmark S. Administration of different Lactobacillus strains in fermented oatmeal soup: in vivo colonization of human intestinal mucosa and effect on the indigenous flora. Appl Environ Microbiol. 1993;59(1):15-20. Accessed January 17, 2025. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC202048/
CrossRef - Adlerberth I, Ahrne S, Johansson ML, Molin G, Hanson LA, Wold AE. A mannose-specific adherence mechanism in Lactobacillus plantarum conferring binding to the human colonic cell line HT-29. Appl Environ Microbiol. 1996;62(7):2244-2251. doi:10.1128/aem.62.7.2244-2251.1996
CrossRef - Molenaar D, Bringel F, Schuren FH, de Vos WM, Siezen RJ, Kleerebezem M. Exploring Lactobacillus plantarum Genome Diversity by Using Microarrays. J Bacteriol. 2005;187(17):6119-6127. doi:10.1128/JB.187.17.6119-6127.2005
CrossRef - Junnarkar M, Pawar S, Gaikwad S, Mandal A, Jass J, Nawani N. Probiotic potential of lactic acid bacteria from fresh vegetables : Application in food preservation. Indian Journal of Experimental Biology. 2019;57:825-838. Accessed January 23, 2025. https://urn.kb.se/resolve?urn=urn:nbn:se:his:diva-17980
- Tripathi N, Sapra A. Gram Staining. In: StatPearls. StatPearls Publishing; 2024. Accessed July 22, 2024. http://www.ncbi.nlm.nih.gov/books/NBK562156/
- Paray AA, Singh M, Mir MA, kaur D. Gram Staining: A Brief Review. International Journal of Research and Review. 2023;10:336-341. doi:10.52403/ijrr.20230934
CrossRef - roy setu, Sawrav M, Das S, et al. Isolation and Biochemical Characterization of Cellulase Producing Goat Rumen Bacteria. In: ; 2022:93-101. doi:10.21467/proceedings.123.12
CrossRef - Khushboo, Karnwal A, Malik T. Characterization and selection of probiotic lactic acid bacteria from different dietary sources for development of functional foods. Front Microbiol. 2023;14:1170725. doi:10.3389/fmicb.2023.1170725
CrossRef - Hutahaean AJN, Silalahi J, Suryanto D, Satria D. Characterisation of Lactic Acid Bacteria from Dengke Naniura of Common Carp (Cyprinus carpio) with α-Glucosidase Inhibitory Activity. Open Access Maced J Med Sci. 2019;7(22):3794-3798. doi:10.3889/oamjms.2019.506
CrossRef - Lee EHL, Degener JE, Welling GW, Veloo ACM. Evaluation of the Vitek 2 ANC Card for Identification of Clinical Isolates of Anaerobic Bacteria▿. J Clin Microbiol. 2011;49(5):1745-1749. doi:10.1128/JCM.02166-10
CrossRef - Li Y, Gu B, Liu G, et al. MALDI-TOF MS versus VITEK 2 ANC card for identification of anaerobic bacteria. J Thorac Dis. 2014;6(5):517-523. doi:10.3978/j.issn.2072-1439.2014.02.15
- Ajazuddin, Saraf S. Evaluation of physicochemical and phytochemical properties of Safoof-E-Sana, a Unani polyherbal formulation. Pharmacognosy Res. 2010;2(5):318-322. doi:10.4103/0974-8490.72332
CrossRef - González C, Kharissova O, Martínez-Hernández A, Castaño V, Velasco-Santos C. Graphene oxide sheets covalently grafted with keratin obtained from chicken feathers. Digest Journal of Nanomaterials and Biostructures. 2015;8:127-138.
- Kielkopf CL, Bauer W, Urbatsch IL. Bradford Assay for Determining Protein Concentration. Cold Spring Harb Protoc. 2020;2020(4):pdb.prot102269. doi:10.1101/pdb.prot102269
CrossRef - Kim SY, Park SY, Jin JE, et al. Comparing the VITEK 2 ANC card, species-specific PCR, and MALDI-TOF mass spectrometry methods for identification of lactic acid bacteria. J Food Sci. 2022;87(11):5099-5106. doi:10.1111/1750-3841.16343
CrossRef - Catalase Test. ASM.org. Accessed July 22, 2024. https://asm.org:443/Protocols/Catalase-Test-Protocol
- Mahon C, Lehman D, Manuselis G. Textbook of Diagnostic Microbiology, 5th edition. Clinical Research and Leadership Faculty Publications. Published online January 1, 2015. https://hsrc.himmelfarb.gwu.edu/smhs_crl_facpubs/37
- Wheelis M. Principles of Modern Microbiology. Jones & Bartlett Learning; 2008.
Abbreviations
% TA : % Titratable Acidity; ANOVA : Analysis of Variance; BSA : Bovine Serum Albumin; CFU – Colony Forming Unit; Doubling Time: Dt; E. coli : Escherichia coli; FOS : Fructooligosaccharide; H2O2 : Hydrogen Peroxide; L. plantarum : Lactobacillus plantarum; MEM : Minimal Essential Medium; MRS : de Man, Rogosa, and Sharpe; NaOH : Sodium Hydroxide; ns : Non-significant ; OD : Optical Density; SCFAs – Short-Chain Fatty Acids; SEM : Standard Error of the Mean; TSI : Triple Sugar Iron; USP : United States Pharmacopeia; Zone of Inhibition : ZOI

This work is licensed under a Creative Commons Attribution 4.0 International License.






