

Introduction
As people are living longer,1 the prevalence of chronic conditions, including diabetes mellitus, cancer and cardiovascular disease is increasing.2 As we age, our immune system alters and older adults, particularly those who are immunocompromised, are more susceptible to infection.3 Evidence suggests that good nutrition plays a key role in promoting healthy aging.4 It has been suggested that incorporating the right foods and nutrients in the diet can help reduce the incidence of age-related diseases5 ,and the need for pharmaceutical interventions.6,7 Furthermore, incorporation of whole foods may be more beneficial than food supplements, as often isolated compounds lose bioactivity.8
Fruit and dairy are essential dietary components.9,10 Fruits provide vitamins and minerals, and are also a rich source of phytochemicals that act as antioxidants,11 which can help prevent chronic illnesses and reduce disease-related mortality.12 Dairy products (milk, yogurt and cheese) are a good source of calcium,13 which helps prevent bone loss in the elderly14, and are also a source of bioactive compounds that have many associated health benefits.15,16 Individually, cheese and fruit consumption have been associated with a reduced risk of type 2 diabetes. 17,18 The antimicrobial potential of cheese has also been well documented19–23 and few studies have reported on the antimicrobial properties of fruit including berries, blackcurrants, grapes and peppers.24–26 Despite the potential health benefits, the Irish Longitudinal Study on Aging reported that 75% of older adults do not consume the recommended dietary allowance (RDA) of fruit and vegetables, and 70% of the older adult population consume less than the recommend daily serving of dairy.27
Combining fruit and dairy products has the potential to promote synergistic health benefits.28 Food synergy relates to combined food ingredients or food matrices that demonstrate improved health benefits beyond their basic nutritional composition, compared to the individual foods or ingredients.29 Combining foods or food ingredients can also have antagonistic effects,30 whereby the sum of the effects is less than that from the individual components.31 Previous studies have demonstrated that fortifying dairy products with herbs and fruit, has the potential to improve the antioxidant, antidiabetic and antimicrobial properties of the dairy food. Al-Otaibi et al., 32 reported improved antioxidant properties for mold-ripened cheese following fortification with date palm fruit, while Apostolidis et al., 33 reported the ability of cranberry enriched cheese to inhibit key enzymes relevant to carbohydrate metabolism. In addition, Khalifa and Wahdan., 34 demonstrated that the addition of cranberry fruit extract to soft white Domiati cheese significantly improved the antimicrobial properties of the cheese and reduced microbial growth during storage.
Studies have highlighted the potential health benefits of combining dairy and fruit, however; to the best of our knowledge, the bioactive potential of cheese-fruit combinations following in vitro digestion has not been reported. Rashidinejad et al., 35 investigated the effects of combining a full-fat hard cheese matrix with green tea catechins on the bioactive properties of cheese, following in vitro digestion, and confirmed that the addition of the green tea extracts increased the antioxidant properties of the cheese.
Novel cheese-fruit products could help address the low dietary intake of dairy and fruit in the older adult population.27 In this study, selected cheese matrices were combined with different fruit and then digested using a simulated gastrointestinal in vitro digestion (SGID) model representative of older adults. The digestates were then assessed to determine if the combinations altered the potential antioxidant, α-glucosidase inhibitory and immunomodulatory properties of the cheese. The antimicrobial properties of the cheese-fruit combination were also investigated. Cheese-fruit combinations with demonstrated synergistic effects could be considered as functional foods for older adults with potential to promote healthy aging.
Materials and Methods
All chemicals were purchased from Merck (Sigma-Aldrich, Ireland), unless otherwise stated. The Jurkat cell line was purchased from the European Collection of Authenticated Cell Cultures (ECACC, UK). Bacterial cultures included Escherichia coli (#DSM3008; DSMZ, Germany), Listeria monocytogenes36 and Group B Streptococcus (GBS)37. Antimicrobial agents included Gentamicin (10 µg) and Penicillin G (10U) (Oxoid, Thermofisher scientific, Ireland). Ringers tablets, brain heart infusion (BHI) broth and BHI agar were purchased from LAB M, UK.
Sample Selection and Preparation
Cheese and fruit products were purchased from local supermarkets (within one season) and included reduced fat red cheddar (RFRC), feta, goat’s cheese, goji berries, red peppers, and blackberries. Cheese products were selected to represent cheese with different textures and fat content, as it has previously been reported that cheese varying in texture and fat content have different digestibility rates.38 The red cheddar was a reduced fat, hard cheese made from cows’ milk; the feta was a full-fat soft cheese made from sheep and goat’s milk, and the goat’s cheese was a medium-fat soft cheese made from goat’s milk. Three types of fruit including goji berries, red pepper and blackberries were selected as they are all known to be good sources of antioxidant compounds and are associated with a range of bioactive properties.39–42 Food combinations were prepared by mixing 18 g of cheese with 2 g of fruit in 20 mL of distilled deionized H2O for 2 min. The composition of each cheese, as identified by the manufacturer, is summarized in Table 1.
Table 1: Composition of Cheese Products Purchased from Local Supermarkets.
| Cheese type per 100g | Energy(Kcal) | Fat(g) | Saturates(g) | Protein(g) | Salt(g) |
| Reduced fat red cheddar | 302.0 | 22.0 | 14.0 | 28.0 | 2.0 |
| Feta | 279.0 | 23.0 | 17.1 | 16.9 | 1.9 |
| Goat’s cheese | 158.0 | 12.0 | 8.0 | 9.5 | 1.3 |
In Vitro Digestion Procedure
To investigate and compare the potential bioactive properties of the cheese-fruit combinations, each combination (10 % w/w) was subjected to simulated gastrointestinal in vitro digestion (SGID) using a method described by Plante et al.,43 modified from the SGID model described by Minekus et al.,44 to reflect some of the physiological changes associated with an older adult. Briefly, 5 g of sample was homogenized with 3.5 mL of simulated salivary fluid, 25 µL CaCl2 (0.3M) and water to a final volume of 10 mL, and then incubated at 37oC in a shaking incubator for 2 min. Following this, the gastric phase was simulated by mixing 10 mL of oral digestate with 7.5 mL simulated gastric fluid (pH 3), 750 U mL-1 porcine pepsin, 5 µL CaCl2 (0.3M), 0.2 mL HCL (1M) and water to achieve a final volume of 20 mL, and incubated at 37oC in a shaking incubator for 2 hr. The final phase of digestion, the intestinal phase, was simulated by mixing 20 mL of gastric digestate with 11 mL of simulated intestinal fluid (pH 6.5), 5.0 mL of pancreatin solution (800 U mL-1), 2.0 mL (10mM) bile, 40 µL of CaCl2 (0.3M), 0.15 mL NaOH (1M) and water to achieve a final volume of 40 mL, and incubated at 37oC in a shaking incubator for 2 hr.
Antioxidant Activity
Chemical-based in vitro antioxidant assays are useful screening tools as they are low cost, allow for high-throughput, and yield an index value that helps to compare the potential antioxidant properties of different compounds and products.45 The principles of the assays can vary and, therefore, it is recommended that more than one assay is used to assess antioxidant activities.45
DPPH (2,2-di-phenyl-1-picryl hydrazyl) Radical Scavenging Activity
Cheese digestates were investigated for their ability to scavenge the DPPH free radical using a previously described method,46 with modifications43 ;1ml of sample was mixed with 3 mL of 60 μM DPPH/methanol solution. Color blanks were prepared with 1 mL of digestate and 3 mL of methanol, and controls consisted of 1 mL of methanol to 3 mL of 60 μM DPPH/methanol solution. %DPPH inhibition was calculated against the control and compared to a Trolox standard curve (0.04 – 0.4 µM).
![]()
Ferric Reducing Antioxidant Power
Reducing power of cheese digestates was assessed according to Benzie and Strain.,47 with modifications43; Samples (1 mL) were combined with 2 mL of FRAP reagent and, color blanks were prepared with H2O in place of FRAP reagent. A mix of FRAP reagent (2 mL) and distilled deionized H2O (1 mL) was used as a blank. Results were expressed as micromole of ferrous per kg.fw of cheese sample (µmol Fe2+/kg.fw).
Total Phenolic Content
TPC of cheese digestates was measured by Folin-Ciocalteu method,48 with modifications.43 Samples (50 µL) were added to Folin-Ciocalteau solution (250 µL) and incubated for 4 min. Then, , 500 µL of 2 % (w/v) Na2CO3 and 4.2 mL of H2O were added. Color blanks consisted of 50 µL of sample, 4.45 mL of H2O and 500 µL of 2 % (w/v) Na2CO3. After 120 min at20°C , absorbance was measured at 765 nm versus a water blank. TPC was determined against a standard curve of gallic acid (0 – 50 mg/mL) and expressed as mg gallic acid equivalents (GAE) per 100 g of fresh sample (mg GAE/100g.fw).
α-Glucosidase Inhibition
The ability of digestates to inhibit α-glucosidase, an enzyme responsible for the digestion of complex carbohydrates in vivo,49 was investigated using a previously described method.33 Sample (50 μL) was mixed with 0.1 M phosphate buffer (100 μL, pH 6.9) containing α-glucosidase solution (1.0 U/mL) in a 96-well plate at 25 °C for 10 min.Then , 50 μL of substrate (5 mM p-nitrophenyl-α-D-glucopyranoside solution in 0.1 M phosphate buffer, pH 6.9) was added to each well. Color blanks consisted of 50 μL of 0.1 M phosphate buffer (pH 6.9) in place of enzyme, and the blank was a mixture of buffer and substrate. Reactions were incubated at 25 °C for 5 min. Absorbance was recorded at 405 nm by a microplate reader (Varioskan Flash microplate reader, ThermoScientific). Results were compared to the control which had 50 μL of buffer solution in place of the extract. The α-glucosidase inhibitory potential was expressed as % inhibition and calculated as follows:
![]()
Immunomodulatory Activity
Cytotoxicity was investigated using the MTT assay previously described by Gabrani et al., 50 with modifications. Digestates were analyzed at concentrations of 0-5 % (v/v). Controls consisted of no digestate and media only used as a blank. Cell viability was calculated as follows:
![]()
Non-toxic concentration of 0.5 % (v/v) was selected to analyze the immunomodulatory properties, with >80 % average cell viability. Immunomodulatory potential was investigated using a previously described method with modifications.51 Digestates were examined for their potential to modulate TNF-α levels in stimulated Jurkat T cells that were grown in T25 culture flasks (2 x 105 cells per mL) with reduced serum media (RPMI/FBS 5 %). Cells were treated with concanavalin A (conA, 50 μg/mL), and incubated in a 96 well plate (100 µL per well) with 0.5 % (v/v) sample at 37 ˚C in a 5 % CO2 atmosphere for 24 h. Controls included; (i) Media (5 % RPMI/FBS), (ii) Cells and media (5 % RPMI/FBS), and (iii) Cells treated with conA and media (5 % RPMI/FBS). Plates were then centrifuged (106 g x 10 min), and supernatants collected and stored at -80 ºC until analyzed. TNF-alpha was measured by ELISA (Human TNF-alpha, R & D systems), and absorbance was read at 450 nm on a microplate reader (Varioskan Flash microplate reader, ThermoScientific).TNF-alpha production was determined using online analysis software (elisaanalysis.com) and expressed as pg/mL.
Antimicrobial Activity
Antimicrobial activity of digestates was screened using a disk diffusion assay according to a method described by Meira et al.52 Bacterial cultures of Escherichia coli, Listeria monocytogenes and Group B Streptococcus were diluted in Ringers buffer to prepare suspensions at 108 cfu/mL. Cultures were inoculated onto BHI agar plates using a sterile swab. Sterile discs were placed aseptically onto the surface of the plates, 15 μL of digestate was added to the discs and incubated at 37 °C for 24 h to identify zones of inhibition. Antibiotic discs used as positive controls included Penicillin G (10 U) (for GBS and L. monocytogenes cultures) and Gentamicin (10 µg) (for E. coli cultures). Negative controls consisted of sterile H2O. Interpretation of growth inhibition was based on measurement (mm) of zones of clearing.
Statistical Analysis
All data was summarized with a mean ± standard deviation of at least three independent experiments. Statistical analysis was carried out using the IBM Statistical Package for Social Sciences (SPSS v.26). All the data satisfied the conditions of normality and homogeneity of variance, hence a one-way analysis of variance (ANOVA) was used to compare differences in the bioactivity between samples obtained following in vitro digestion. Controlling for multiple comparisons, the Dunnett’s post-hoc test was used to evaluate mean changes between each combination group and the control group. All statistical test results were interpreted using a 5% level of significance.
Results
Antioxidant Activity
Antioxidant potential of individual fruits (goji berries, red pepper, blackberries), plain cheese (feta, goats, RFRC) and, cheese-fruit combinations were investigated by measuring radical scavenging properties and reducing potential of the digested food products. Radical scavenging properties of individual fruits and plain soft cheese (feta or goats) were high (≥83% DPPH inhibition, Figure 1a), and this was not further enhanced when the cheese and fruit were combined (p > 0.05, Figure 1a, Table 2). However, the radical scavenging properties of hard cheese (RFRC) significantly increased, when combined with all three fruits (p < 0.05, Figure 1a, Table 2).
Reducing power of individual fruits significantly differed (p < 0.05) from each other with goji berries demonstrating greatest FRAP (762.4 ± 15.2 µmol Fe2+/kg.fw, Figure 1b). Goat’s cheese had significantly higher (p < 0.05) reducing potential (718.0 ± 13.7 µmol Fe2+/kg.fw) compared to the feta and RFRC (278.9 ± 4.9, 345.8 ± 2.7 µmol Fe2+/kg.fw, respectively, Figure 1b). However, FRAP of all cheese significantly increased (p < 0.05) when combined with the fruits, compared to the plain cheese, and also compared to the fruit alone (Figure 1b, Table 2). TPC of individual fruits was significantly different (p < 0.05), with goji berries demonstrating greatest levels (710.6 mg GAE/100g.fw, Figure 1c). Regarding plain cheese, RFRC had significantly higher TPC levels (p < 0.05, 656.4 mg GAE/100g.fw) compared to feta and goat’s cheese (379.5, 312.4 mg GAE/100g.fw, respectively, Figure 1c). The majority of cheese samples (89%) had significantly (p < 0.05) lower TPC levels following the addition of fruit (Figure 1c).
 |
Figure 1: Antioxidant Activity of Fruit, Plain Cheese and Cheese-Fruit Combinations following In Vitro Digestion. |
α-Glucosidase Inhibition
All three fruit digestates (goji berries, red pepper, and blackberries) demonstrated high α-glucosidase inhibition (≥ 99 % inhibition; Figure 2). Plain soft cheese (Feta and goats) had significantly greater (p < 0.05) α-glucosidase inhibitory properties (96.7 ± 0.9, 98.5 ± 1.3 %inhibition, respectively) compared to the RFRC hard cheese product (17.2 ± 1.1 %inhibition, Figure 2). The α-glucosidase inhibitory potential of feta cheese significantly decreased (p < 0.05), when combined with all three fruits, although the cheese-fruit combinations still retained a high level of inhibition (83 ± 2.5 to 93 ± 0.3 %inhibition; Figure 2, Table 2). Plain goat’s cheese and its combinations were comparable in enzyme inhibitory activity (≥ 98 % inhibition) with no significant difference observed (p > 0.05, Table 2). RFRC had low inhibitory potential which significantly increased, when combined with all three fruits (p < 0.05, Figure 2, Table 2).
 |
Figure 2: α-glucosidase Inhibitory Potential of Fruit, Plain Cheese and Cheese-Fruit Combinations following In Vitro Digestion. |
Immunomodulatory Properties
The immunomodulatory potential of the digestates was investigated by examining their ability to alter cytokine levels in Jurkat T lymphocytes. Concanavalin-A (conA) was used as a positive control as it is commonly used as a stimulant of T-cell activation in vitro and has been shown to stimulate cytokine production in leukocytes.51 All digested fruit resulted in a significant increase in TNF-α levels, compared to the positive control (p < 0.05, Figure 3).
Feta cheese was the only plain cheese that significantly increased TNF-α production compared to the positive control (p < 0.05, Figure 3). However, when comparing TNF-α secretion of the fortified cheese relative to the plain cheese, TNF-α secretion significantly reduced for feta cheese, when combined with all three fruits (Goji berries, red peppers and blackberries) (p < 0.05, Table 2), and there was no significant difference in TNF-α secretion observed when goat’s cheese or RFRC cheese was combined with the fruit (p > 0.05, Table 2).
Antimicrobial Properties
None of the digestates examined in this study (individual cheese or fruit, cheese-fruit combinations) displayed antimicrobial properties against E. coli or L. monocytogenes (data not shown). However, some digestates did inhibit the growth of GBS. Penicillin G remains the first-line treatment for invasive GBS disease in adults53 and thus, was used as a positive control in this study. Penicillin G inhibited the growth of GBS with a strong zone of inhibition (37.0 ± 1.5 mm). Regarding individual fruits, both red pepper and blackberries inhibited the growth of the GBS strain, with comparable zones of inhibition (7.7 ± 0.5, 7.0 ± 1.0 mm, respectively), however no inhibition was observed for the digested goji berries. Goat’s cheese was the only plain cheese that demonstrated antimicrobial activity against GBS (8.6 ± 1.5 mm), and this was not further enhanced when the cheese and fruit were combined, with no significant difference observed (p > 0.05, Table 2). Plain feta cheese did not display antimicrobial activity against GBS, but when combined with red pepper and blackberries, the cheese-fruit combination exerted antimicrobial properties (p < 0.05, Table 2). Plain RFRC and its combinations did not display antimicrobial properties against GBS.
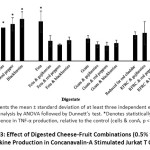 |
Figure 3: Effect of Digested Cheese-Fruit Combinations (0.5% v/v) on Cytokine Production in Concanavalin-A Stimulated Jurkat T Cells. |
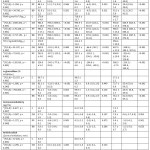 |
Table 2: Comparison of the Bioactive Properties of Cheese-Fruit Combinations, Comparing Plain Cheese (Control) with Cheese-Fruit Combinations. |
Discussion
Across Europe, the intake of dairy products among older adults is lower than recommended54 and less than a quarter of older adults eat the recommended servings of fruit and vegetables per day.55 One potential strategy to address this issue, is the use of food combinations to generate novel products; however,it is of interest to first identify the most effective combinations to ensure synergistic, rather than antagonistic effects.56
In this study, different cheese matrices were combined with fruit and the bioactive properties of the combinations were evaluated following in vitro digestion. In vitro antioxidant assays are frequently used to investigate the antioxidant potential of foods,57 and as screening tools are less expensive, complex and labour intensive.45 The reducing antioxidant properties of all cheese analyzed significantly improved when combined with the fruit (p < 0.05). Combining reduced fat red cheddar (RFRC) with the fruit also significantly improved the radical scavenging properties of the cheese (p < 0.05). Lee et al., 58 also reported that the radical scavenging properties of cheddar cheese can be improved when combined with plant extracts. TPC levels were significantly lower for the majority of the cheese matrices when combined with fruit (p < 0.05), suggesting that phenols may not be solely responsible for the antioxidant properties observed. McDougall et al., 59 observed a similar effect when a dairy matrix (ice-cream) was combined with raspberries.59 A decrease in TPC levels may be due to reduced enzyme secretion in an older adult digestive system and the impact this has on protein-polyphenol interactions. While some interactions can prevent phenolic compounds from being degraded, others may hinder their release from a food matrix.60
The incidence of type II diabetes increases with age.61 One antidiabetic strategy is to inhibit key enzymes relevant to glucose metabolism.62 In this study, the α-glucosidase inhibitory potential of cheese-fruit combinations were investigated, and all digestates displayed high inhibitory potential (≥58% inhibition). Similar to Apostolidis et al.,33 this study demonstrated that cheese enriched with berries inhibited α-glucosidase, however, results from this study also demonstrated that this property was retained following in vitro digestion. Berries have been shown to be effective α-glucosidase inhibitors, largely due to their tannin content.63 RFRC combined with all fruit significantly improved the enzyme inhibitory properties of the cheese (p < 0.05). However, the α-glucosidase inhibitory potential of feta cheese significantly decreased when combined with the fruit, suggesting potential antagonistic effects (p < 0.05). The antagonistic effects observed may be due to specific protein-polyphenol interactions in the feta-fruit combinations.64 Ni et al., 65 observed similar effects when combining a soft dairy matrix (yogurt) with berries, which reduced the antidiabetic properties of the yogurt.
Aged-related changes in the immune system include cytokine dysregulation.66 Tumor necrosis factor-α (TNF-α) is an important immune regulator67 that has both anti-inflammatory68 and pro-inflammatory properties.69 Abnormal levels of TNF-α are associated with conditions such as rheumatoid arthritis, Crohn’s disease, atherosclerosis, psoriasis, sepsis, diabetes, and obesity.69 Aging is associated with elevated levels of TNF-α,68 thus food-based strategies that regulate levels of this important immunomodulatory agent could help modulate the immune response in older adults. The immunomodulatory properties of feta cheese were significantly altered following enrichment with fruit, with a reduction in TNF-α secretion observed (p < 0.05). This may be due to a combination of bioactive compounds in the cheese-fruit mixture, including anti-inflammatory compounds that can be found in fruit40–42 and conjugated linoleic acids (CLA) in cheese. Cheese made from sheep’s milk is naturally rich in CLA and has been associated with anti-inflammatory properties.70 López-García et al., 71 investigated the anti-inflammatory effects of a sterol enriched milk-based fruit beverage and found that the beverage demonstrated moderate anti-inflammatory effects and suggested that a combination of bioactive compounds may be responsible for the effect observed.
Aging is also associated with an increased susceptibility to infection.3 Group B Streptococci (GBS) infections are increasing in older adult populations,53,72,73 with serotype V being the most prevalent serogroup associated with invasive disease in adults.74 The gastrointestinal tract is a reservoir for GBS75 and colonization of the gut is considered a first crucial step in the progression of infections.76 Antimicrobial peptides have been previously identified in cheese20,21 and fruit.77,78 Interestingly, feta cheese was not active against GBS, until combined with fruit suggesting potential synergistic effects.79,80 Digested hard cheese (cheddar) displayed no bactericidal effect, even when combined with the fruit. Fang et al., 38 confirmed that soft cheese matrices are easily disrupted during gastric digestion, with a fast release of peptides compared to harder cheese matrices, which may be linked to the lack of antimicrobial activity of the RFRC product. Further studies are required to confirm the antimicrobial properties of the cheese-fruit combinations against other GBS strains and serotypes, but these preliminary studies suggest that a diet-based strategy to reduce GBS colonization in vivo and limit GBS infections in older adults warrants further investigation.53
It is important to acknowledge that this study was based on in vitro investigations and further studies would be necessary to confirm in vivo effects. In addition, future work could identify the compounds responsible for the activities observed. The current study adds to existing research that supports combining cheese with other food matrices, such as fruit, to enhance the bioactive properties of cheese. Understanding the potential synergistic and/or antagonistic effects that can occur when dairy and fruit matrices are combined could help aid the design of novel functional foods for older adults.
Conclusion
To date, studies reporting on the synergistic effects observed when cheese and fruit are combined are limited, and no studies to date have examined these effects following in vitro digestion with an older adult gut model. Certain cheese products combined with fruit significantly improved the antioxidant, α-glucosidase inhibitory, antimicrobial, and immunomodulatory properties of the cheese. In particular, the antioxidant and α-glucosidase inhibitory potential of reduced fat red cheddar significantly improved when combined with fruit. Cheddar cheese is a product that is well received by older adults, who also have a preference towards reduced fat products and low-calorie intake. Novel RFRC-fruit combinations could be an attractive functional food option for this cohort.
Acknowledgements
The authors would like to thank Munster Technological University Ireland, for funding this research.
Funding Source
This work was supported by Munster Technological University (Formerly, Cork Institute of technology), Ireland under the RÍSAM Scholarship scheme.
Conflict of Interest
The authors declare no conflict of interest.
References
- Lunenfeld B., Stratton P. The clinical consequences of an ageing world and preventive strategies. Best Pract Res Clin Obstet Gynaecol. 2013;27(5):643-659. doi:10.1016/J.BPOBGYN.2013.02.005.
CrossRef - Jaul E., Barron J. Age-Related Diseases and Clinical and Public Health Implications for the 85 Years Old and Over Population. Front public Heal. 2017;5:335. doi:10.3389/fpubh.2017.00335.
CrossRef - Rescigno T., Micolucci L., Tecce MF., Capasso A. Bioactive Nutrients and Nutrigenomics in Age-Related Diseases. Molecules. 2017;22(1):105. doi:10.3390/molecules22010105.
CrossRef - Shlisky J., Bloom DE., Beaudreault AR., et al. Nutritional considerations for healthy aging and reduction in age-related chronic disease. Adv Nutr An Int Rev J. 2017;8(1):17-26. doi:10.3945/an.116.013474.
CrossRef - Everitt A V., Hilmer SN., Brand-Miller JC., et al. Dietary approaches that delay age-related diseases. Clin Interv Aging. 2006;1(1):11-31. doi:10.2147/ciia.2006.1.1.11.
CrossRef - Eussen S., Klungel O., Garssen J., et al. Support of drug therapy using functional foods and dietary supplements: Focus on statin therapy. Br J Nutr. 2010;103(9):1260-1277. doi:10.1017/S0007114509993230.
CrossRef - Scolaro B., Soo Jin Kim H., de Castro IA. Bioactive compounds as an alternative for drug co-therapy: Overcoming challenges in cardiovascular disease prevention. Crit Rev Food Sci Nutr. 2018;58(6):958-971. doi:10.1080/10408398.2016.1235546.
CrossRef - Liu RH. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am J Clin Nutr. 2003;78(3):517S-520S. doi:10.1093/ajcn/78.3.517S.
CrossRef - Food Safety Authority of Ireland. Scientific Recommendations for Healthy Eating Guidelines in Ireland. FSAI;2011:1-92. https://www.fsai.ie/WorkArea/DownloadAsset.aspx?id=16765. Accessed Janurary 11, 2019.
- Food and Agriculture Organization of the United Nations. Food-based dietary guidelines – Ireland. FAO;2012. http://www.fao.org/nutrition/education/food-dietary-guidelines/regions/countries/ireland/en/. Accessed January 11, 2019.
- Slavin JL., Lloyd B. Health Benefits of Fruits and Vegetables. Adv Nutr. 2012;3(4):506-516. doi:10.3945/an.112.002154.
CrossRef - Kadell A., Nicklett E. Fruit and vegetable intake among older adults. Maturitas. 2013;75(4):305-312. doi:10.1016/j.maturitas.2013.05.005.
CrossRef - Thorning TK., Raben A., Tholstrup T., Soedamah-Muthu SS., Givens I., Astrup A. Milk and dairy products: Good or bad for human health? An assessment of the totality of scientific evidence. Food Nutr Res. 2016;60:32527. doi:10.3402/fnr.v60.32527.
CrossRef - Rene R. Dairy products, yogurts, and bone health. Am J Clin Nutr. 2014;99(5):1256S-1262S. doi:10.3945/ajcn.113.073056.
CrossRef - Korhonen H. Milk-derived bioactive peptides: From science to applications. J Funct Foods. 2009;1(2):177-187. doi:10.1016/j.jff.2009.01.007.
CrossRef - Park YW., Nam MS. Bioactive peptides in milk and dairy products: a review. Korean J food Sci Anim Resour. 2015;35(6):831-840. doi:10.5851/kosfa.2015.35.6.831.
CrossRef - Sluijs I., Forouhi G N., Beulens WJ J., et al. The amount and type of dairy product intake and incident type 2 diabetes: Results from the EPIC-InterAct Study. Am J Clin Nutr. 2012;96(2):382-390. doi:10.3945/ajcn.111.021907.
CrossRef - Wang P-Y., Fang J-C., Gao Z-H., Zhang C., Xie S-Y. Higher intake of fruits, vegetables or their fiber reduces the risk of type 2 diabetes: A meta-analysis. J Diabetes Investig. 2016;7(1):56-69. doi:10.1111/jdi.12376.
CrossRef - Rizzello CG., Losito I., Gobbetti M., Carbonara T., De Bari MD., Zambonin PG. Antibacterial activities of peptides from the water-soluble extracts of Italian cheese varieties. J Dairy Sci. 2005;88(7):2348-2360. doi:10.3168/jds.S0022-0302(05)72913-1.
CrossRef - Losito I., Carbonara T., De Bari MD., et al. Identification of peptides in antimicrobial fractions of cheese extracts by electrospray ionization ion trap mass spectrometry coupled to a two-dimensional liquid chromatographic separation. Rapid Commun Mass Spectrom. 2006;20(3):447-455. doi:10.1002/rcm.2323.
CrossRef - Pritchard SR., Phillips M., Kailasapathy K. Identification of bioactive peptides in commercial Cheddar cheese. Food Res Int. 2010;43(5):1545-1548. doi:10.1016/j.foodres.2010.03.007.
CrossRef - Mohanty DP., Mohapatra S., Misra S., Sahu PS. Milk derived bioactive peptides and their impact on human health – A review. Saudi J Biol Sci. 2016;23(5):577-583. doi:10.1016/j.sjbs.2015.06.005.
CrossRef - Rahmawati IS., Suntornsuk W. Effects of Fermentation and Storage on Bioactive Activities in Milks and Yoghurts. Procedia Chem. 2016;18:53-62. doi:10.1016/j.proche.2016.01.010.
CrossRef - Cavanagh HMA., Hipwell M., Wilkinson JM. Antibacterial activity of berry fruits used for culinary purposes. J Med Food. 2003;6(1):57-61. doi:10.1089/109662003765184750.
CrossRef - O’Mahony R. The antibacterial properties of dietary fruit. In: Watson RR., Preedy VR. Bioactive foods in promoting health. Elsevier Inc; 2010:147-166 https://www.elsevier.com/books/bioactive-foods-in-promoting-health/watson/978-0-12-374628-3. Accessed April 30, 2019.
- Meneguetti BT., Machado L dos S., Oshiro KGN., Nogueira ML., Carvalho CME., Franco OL. Antimicrobial peptides from fruits and their potential use as biotechnological Tools-A review and outlook. Front Microbiol. 2016;7:2136. doi:10.3389/fmicb.2016.02136.
CrossRef - Mcgarrigle C., Donoghue O., Scarlett S., et al. Health and Wellbeing: Active Ageing for Older Adults in Ireland. Evidence from The Irish Longitudinal Study on Ageing. 2017:1-218. https://tilda.tcd.ie/publications/reports/pdf/w3-key-findings-report/TILDA%20Wave%203%20Key%20Findings%20report.pdf. Accessed March 10, 2020.
- Fernandez MA., Marette A.. Potential Health Benefits of Combining Yogurt and Fruits Based on Their Probiotic and Prebiotic Properties. Adv Nutr An Int Rev J. 2017;8(1):155S-164S. doi:10.3945/an.115.011114.
CrossRef - Jacobs DR., Gross MD., Tapsell LC., Tapsell LC. Food synergy: an operational concept for understanding nutrition. Am J Clin Nutr. 2009;89(5):1543S-1548S. doi:10.3945/ajcn.2009.26736B.
CrossRef - Maurya VK. Factors influencing the absorption of vitamin D in GIT : an overview. J Food Sci Technol. 2017;54(12):3753-3765. doi:10.1007/s13197-017-2840-0.
CrossRef - Wang S., Meckling KA., Marcone MF., Kakuda Y., Tsao R. Synergistic, additive, and antagonistic effects of food mixtures on total antioxidant capacities. J Agric Food Chem. 2011;59(3):960-968. doi:10.1021/jf1040977.
CrossRef - Al-Otaibi MM., Haddadin JS., Haddadin MSY. Mold-ripened soft cheeses fortified with date palm fruit product as functional dairy products. Pakistan J Biol Sci. 2016;19(1):11-25. doi:10.3923/pjbs.2016.11.25.
CrossRef - Apostolidis E., Kwon YI., Shetty K. Inhibitory potential of herb, fruit, and fungal-enriched cheese against key enzymes linked to type 2 diabetes and hypertension. Innov Food Sci Emerg Technol. 2007;8(1):46-54. doi:10.1016/j.ifset.2006.06.001.
CrossRef - Khalifa SA., Wahdan KM. Improving the quality characteristics of white soft cheese using cranberry (Vaccinium macrocarpon) fruit extract. Int Food Res J. 2015;22(6):2203-2211. http://www.ifrj.upm.edu.my/22%20(06)%202015/(4).pdf
- Rashidinejad A., Birch EJ., Everett DW. Antioxidant activity and recovery of green tea catechins in full-fat cheese following gastrointestinal simulated digestion. J Food Compos Anal. 2016;48:13-24. doi:10.1016/j.jfca.2016.02.004.
CrossRef - Linnan MJ., Mascola L., Lou XD., et al. Epidemic listeriosis associated with Mexican-style cheese. N Engl J Med. 1988;319(13):823-828. doi:10.1056/NEJM198809293191303.
CrossRef - Hayes K., Cotter L., Barry L., O’Halloran F. Emergence of the L phenotype in Group B Streptococci in the South of Ireland. Epidemiol Infect. 2017;145(16):3535-3542. doi:10.1017/S0950268817002461.
CrossRef - Fang X., Rioux LE., Labrie S., Turgeon SL. Commercial cheeses with different texture have different disintegration and protein/peptide release rates during simulated in vitro digestion. Int Dairy J. 2016;56:169-178. doi:10.1016/j.idairyj.2016.01.023.
CrossRef - Tavares L., Figueira I., McDougall GJ., et al. Neuroprotective effects of digested polyphenols from wild blackberry species. Eur J Nutr. 2013;52(1):225-236. doi:10.1007/s00394-012-0307-7.
CrossRef - Chávez-Mendoza C., Sanchez E., Muñoz-Marquez E., Sida-Arreola JP., Flores-Cordova MA. Bioactive compounds and antioxidant activity in different grafted varieties of bell pepper. Antioxidants. 2015;4(2):427-446. doi:10.3390/antiox4020427.
CrossRef - Skrovankova S., Sumczynski D., Mlcek J., Jurikova T., Sochor J. Bioactive Compounds and Antioxidant Activity in Different Types of Berries. Int J Mol Sci. 2015;16(10):24673-24706. doi:10.3390/ijms161024673.
CrossRef - Vulić JJ., Čanadanović-Brunet JM., Ćetković GS., Djilas SM., Tumbas Šaponjac VT., Stajčić SS. Bioactive Compounds and Antioxidant Properties of Goji fruits (Lycium barbarum L.) Cultivated in Serbia. J Am Coll Nutr. 2016;35(8):692-698. doi:10.1080/07315724.2016.1142404.
CrossRef - Plante AM., McCarthy AL., O’Halloran F. Cheese as a functional food for older adults: comparing the bioactive properties of different cheese matrices following simulated gastrointestinal in vitro digestion. Int J Food Sci Nutr. 2020:1-14. doi:10.1080/09637486.2020.1825644.
CrossRef - Minekus M., Alminger M., Alvito P., et al. A standardised static in vitro digestion method suitable for food – an international consensus. Food Funct Food Funct. 2014;5(5):1113-1124. doi:10.1039/c3fo60702j.
CrossRef - López-Alarcón C., Denicola A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal Chim Acta. 2013;763:1-10. doi:10.1016/J.ACA.2012.11.051.
CrossRef - Brand-Williams W., Cuvelier ME., Berset C. Use of a free radical method to evaluate antioxidant activity. LWT – Food Sci Technol. 1995;28(1):25-30. doi:10.1016/S0023-6438(95)80008-5.
CrossRef - Benzie IF., Strain JJ. Ferric reducing/antioxidant power assay: direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Methods Enzymol. 1999;299:15-27. doi: 10.1016/s0076-6879(99)99005-5.
CrossRef - Singleton VL., Rossi JA., JR R. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Vitic. 1965;16(3):144-158. https://www.ajevonline.org/content/16/3/144.
CrossRef - Derosa G., Maffioli P. α-Glucosidase inhibitors and their use in clinical practice. Arch Med Sci. 2012;8(5):899-906. doi:10.5114/aoms.2012.31621.
CrossRef - Gabrani R., Jain R., Sharma A., Sarethy I., Dang S., Gupta S. Antiproliferative effect of Solanum nigrum on human leukemic cell lines. Indian J Pharm Sci. 2012;74(5):451-453. doi:10.4103/0250-474X.108421.
CrossRef - McCarthy AL., O’Callaghan YC., Connolly A., Piggott CO., FitzGerald RJ., O’Brien NM. A study of the ability of bioactive extracts from brewers’ spent grain to enhance the antioxidant and immunomodulatory potential of food formulations following in vitro Int J Food Sci Nutr. 2015;66(2):230-235. doi:10.3109/09637486.2014.979314.
CrossRef - Meira SMM., Daroit DJ., Helfer VE., et al. Bioactive peptides in water-soluble extracts of ovine cheeses from Southern Brazil and Uruguay. Food Res Int. 2012;48(1):322-329. doi:10.1016/J.FOODRES.2012.05.009.
CrossRef - High KP., Edwards MS., Baker CJ. Group B Streptococcal Infections in Elderly Adults. Clin Infect Dis. 2005;41(6):839-847. doi:10.1086/432804.
CrossRef - Ribeiro I., Gomes M., Figueiredo D., Lourenço J., Paúl C., Costa E. Dairy Product Intake in Older Adults across Europe Based On the SHARE Database. J Nutr Gerontol Geriatr. 2019;38(3):297-306. doi:10.1080/21551197.2019.1627972.
CrossRef - Baker AH., Wardle J. Sex differences in fruit and vegetable intake in older adults. Appetite. 2003;40(3):269-275. doi:10.1016/S0195-6663(03)00014-X.
CrossRef - Tapsell LC., Neale EP., Satija A., Hu FB. Foods, Nutrients, and Dietary Patterns: Interconnections and Implications for Dietary Guidelines. Adv Nutr. 2016;7(3):445-454. doi:10.3945/an.115.011718.
CrossRef - Huang D., Boxin OU., Prior RL. The chemistry behind antioxidant capacity assays. J Agric Food Chem. 2005;53(6):1841-1856. doi:10.1021/jf030723c.
CrossRef - Lee N-K., Jeewanthi RKC., Park E-H., Paik H-D. Short communication: Physicochemical and antioxidant properties of Cheddar-type cheese fortified with Inula britannica extract. J Dairy Sci. 2015;99(1):83-88. doi:10.3168/jds.2015-9935.
CrossRef - McDougall GJ., Dobson P., Smith P., Blake A., Stewart D. Assessing potential bioavailability of raspberry anthocyanins using an in vitro digestion system. J Agric Food Chem. 2005;53(15):5896-5904. doi:10.1021/jf050131p.
CrossRef - Ribas-Agustí A., Martín-Belloso O., Soliva-Fortuny R., Elez-Martínez P. Food processing strategies to enhance phenolic compounds bioaccessibility and bioavailability in plant-based foods. Crit Rev Food Sci Nutr. 2018;58(15):2531-2548. doi:10.1080/10408398.2017.1331200.
CrossRef - Kalyani RR., Golden SH., Cefalu WT. Diabetes and aging: Unique considerations and goals of care. Diabetes Care. 2017;40(4):440-443. doi:10.2337/dci17-0005.
CrossRef - Kirkman MS., Briscoe VJ., Clark N., et al. Diabetes in older adults. Diabetes Care. 2012;35(12):2650-2664. doi:10.2337/dc12-1801.
CrossRef - Xiao J., Kai G., Yamamoto K., Chen X. Advance in dietary polyphenols as α-Glucosidases inhibitors: a review on structure-activity relationship aspect. Crit Rev Food Sci Nutr. 2013;53(8):818-836. doi:10.1080/10408398.2011.561379.
CrossRef - Yildirim-Elikoglu S., Erdem YK. Interactions between milk proteins and polyphenols: Binding mechanisms, related changes, and the future trends in the dairy industry. Food Rev Int. 2018;34(7):665-697. doi:10.1080/87559129.2017.1377225.
CrossRef - Ni H., Hayes HE., Stead D., Raikos V. Incorporating salal berry (Gaultheria shallon) and blackcurrant (Ribes nigrum) pomace in yogurt for the development of a beverage with antidiabetic properties. Heliyon. 2018;4(10):e00875. doi:10.1016/j.heliyon.2018.e00875.
CrossRef - Brüünsgaard H., Pedersen BK. Age-related inflammatory cytokines and disease. Immunol Allergy Clin North Am. 2003;23(1):15-39. doi:10.1016/S0889-8561(02)00056-5.
CrossRef - Rieckmann P., Tuscano JM., Kehrl JH. Tumor Necrosis Factor-α (TNF-α) and Interleukin-6 (IL-6) in B-Lymphocyte Function. Methods. 1997;11(1):128-132. doi:10.1006/METH.1996.0396.
CrossRef - Bruunsgaard H., Pedersen M., Pedersen BK. Aging and proinflammatory cytokines. Curr Opin Hematol. 2001;8(3):131-136. doi:10.1097/00062752-200105000-00001.
CrossRef - Parameswaran N., Patial S. Tumor necrosis factor-α signaling in macrophages. Crit Rev Eukaryot Gene Expr. 2010;20(2):87-103. doi:10.1615/critreveukargeneexpr.v20.i2.10.
CrossRef - Sofi F., Buccioni A., Cesari F., et al. Effects of a dairy product (pecorino cheese) naturally rich in cis-9, trans-11 conjugated linoleic acid on lipid, inflammatory and haemorheological variables: A dietary intervention study. Nutr Metab Cardiovasc Dis. 2010;20(2):117-124. doi:10.1016/j.numecd.2009.03.004.
CrossRef - López-García G., Cilla A., Barberá R., Alegría A., Recio MC. Effect of a milk-based fruit beverage enriched with plant sterols and/or galactooligosaccharides in a murine chronic colitis model. Foods. 2019;8(4):114. doi:10.3390/foods8040114.
CrossRef - Phares CR., Lynfield R., Farley MM., et al. Epidemiology of Invasive Group B Streptococcal Disease in the United States, 1999-2005. JAMA. 2008;299(17):2056-2065. doi:10.1001/jama.299.17.2056.
CrossRef - Kothari NJ., Morin CA., Glennen A., et al. Invasive group B streptococcal disease in the elderly, Minnesota, USA, 2003-2007. Emerg Infect Dis. 2009;15(8):1279-1281. doi:10.3201/eid1508.081381.
CrossRef - Teatero S., McGeer A., Low DE., et al. Characterization of invasive group B streptococcus strains from the greater Toronto area, Canada. J Clin Microbiol. 2014;52(5):1441-1447. doi:10.1128/JCM.03554-13.
CrossRef - Spencer BL., Deng L., Patras KA., et al. Cas9 contributes to group b streptococcal colonization and disease. Front Microbiol. 2019;10:1930. doi:10.3389/fmicb.2019.01930.
CrossRef - Shabayek S., Spellerberg B. Group B streptococcal colonization, molecular characteristics, and epidemiology. Front Microbiol. 2018;9:437. doi:10.3389/fmicb.2018.00437.
CrossRef - Guzmán-Rodríguez JJ., López-Gómez R., Suárez-Rodríguez LM., et al. Antibacterial activity of defensin PaDef from avocado fruit (Persea americana var. drymifolia) expressed in endothelial cells against escherichia coli and staphylococcus aureus. Biomed Res Int. 2013;2013. doi:10.1155/2013/986273.
CrossRef - Seo HH., Park S., Park S., et al. Overexpression of a defensin enhances resistance to a fruit-specific anthracnose fungus in pepper. PLoS One. 2014;9(5):e97936. doi:10.1371/journal.pone.0097936.
CrossRef - Clare D., Catignani G., Swaisgood H. Biodefense Properties of Milk: The Role of Antimicrobial Proteins and Peptides. Curr Pharm Des. 2005;9(16):1239-1255. doi:10.2174/1381612033454874.
CrossRef - Hayes M., Stanton C., Fitzgerald GF., Ross RP. Putting microbes to work: Diary fermentation, cell factories and bioactive peptides. Part II: Bioactive peptide functions. Biotechnol J. 2007;2(4):435-449. doi:10.1002/biot.200700045.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






