

Introduction
In the US and worldwide, the second most common cause of death is cancer. Dysregulation of the molecular pathways which alters protein expression resulting in uncontrolled cellular proliferation is the most identified underlying mechanism of cancer.1 There has been a significant improvement in anticancer drug discovery using herbal medicine. Poor effectiveness, unwanted side effects, and tumour resistance were associated with drugs that target particular signalling pathways, while drugs that interfere with multiple molecules responsible for tumour growth have more efficacy and targeting different cancer cell line. Multi-targets drugs are considered to be more effective and less susceptible to drug-resistance. 2
Angiogenesis, a process where new vasculature initiated from pre-existing normal vessels. Initiation of angiogenesis starts with localized release of pro and anti-angiogenesis growth factors by endothelial cells (ECs). 3
New blood vessel inhibition properties of angiogenic proteins in cancer cells will induce endothelial cell apoptosis or inhibiting endothelial receptors. Plant derived anti-angiogenesis agents are unlike conventional chemotherapeutic agents. It can be used in lower doses with less cytotoxicity, and over a period of months to years to prevent recurrence from latent micro metastases. 2
Around 100 new natural products have been established for cancer therapy and this type of research is still on going and needed in cancer therapeutic research area. To date only small part of rainforest plant have been studied in cancer research even the potential of this work have been more relevant and pressing from times to times. Zingiber zerumbet from Zingiberaceae family is one of natural product contains a range of different phytochemical groups. This plant is highly consumed in the traditional Asian diet. Zerumbone is one of these phytochemicals. 4 It inhibits cell proliferation in various cancer cell lines with negligible side effects on normal cells and it also exhibits antiangiogenic activities in cancer. 5
Despite numerous researches focused on the effects of Zerumbone as anti-cancer and anti-angiogenesis compound, limited evidence-based systematic review regarding these topics available.
Zerumbone extracted from Zingiber zerumbet is proven to have anti-cancer and anti-angiogenesis properties. The effect of Zerumbone on cancer cells has been established through in vitro. in vivo and ex vivo researches. Receptor and mitochondrial signaling pathway are the first target in the cancer therapy. Studies has shown that the most cytotoxic drugs trigger cell death by targeting cytochrome-c/Apaf-1/caspase-9–dependent multiple proapoptotic and antiapoptotic signaling pathways. Various signaling pathways can modulate apoptosis and angiogenesis by converging on, thereby altering the activity of, common central control points within the apoptotic and angiogenic signaling pathways, which involve related proteins or mediators such as caspases, Bcl-2 proteins,VEGF and chemokines. Zerumbone displays promising anticancer effects through several mechanisms of actions such as cell cycle arrest, apoptosis, modulate multiple molecular targets and inhibition of tumour angiogenesis. 6,7
This systematic review aimed to systematically summarize studies done on Zerumbone, including its mechanisms of actions, therapeutic and prophylactic potential against different cancers and its role as angiogenesis inhibitor.
Methodology
Search Strategy
For identifying articles used in this systematic review, two separated searches were conducted, both searches were performed through four databases which were Science Direct, PubMed, Google Scholar and Scopus. The last search was performed on 11/01/2019. The combinations of the following keywords were used in the first survey: “anticancer, Zerumbone, Zingiber zerumbet”. The keywords for second search included “anti-angiogenesis, anti-angiogenic, anticancer, Zerumbone and Zingiber zerumbet”.
Inclusion and Exclusion Criteria
The inclusion criteria for selecting relevant studies were as follows: article published between 1/01/2008 to 31/12/2018, articles on anticancer properties of Zerumbone extracted from Zingiber zerumbet, articles on antiangiogenic activity of Zerumbone as one of the roles for their anticancer effect, study designs using in vitro, in vivo and ex vivo. Articles which were written in language other than English, and articles that reported the activity of Zerumbone from unknown source were excluded.
Data Extraction
All selected articles that reported the effect of Zerumbone on cancer cells, and role of their angiogenic properties in anticancer effect were further analysed. These following information were gathered: authors, publication year, design of a study, country, sample size, dose and duration, cytotoxic effect on cancer and normal cells and mechanism of its cytotoxic effect.
Article Selection
The articles retrieved from the databases were screened for duplication. After removing the duplicated articles, a reviewer independently screened the retrieved relevant articles by reading the full copy or abstract; inclusion and exclusion criteria were considered at this stage. In addition to that, manual search was conducted from the bibliography section of the selected articles. Finally, supervisor and co-supervisor reviewed the selected articles.
Results
Search Literature
The literature searched conceded a total of 352 records (from January 2008 to December 2018). The numbers of articles obtained from the corresponding databases are as follows; Google scholar (n=142), Science Direct (n=134), PubMed (n=48) and Scopus (n=28). After the online search, duplicate articles were removed. The remaining 146 articles were screened by referring to the title and abstract. 84 articles were excluded due to nonfulfillment of inclusion criteria. From which 62 articles were further evaluated based on the full text, and a total of 19 articles were removed; two were using Zerumbone derivatives, two articles were review articles, six articles did not mention source of Zerumbone from Zingiber zerumbet clearly, one not related to anticancer nor antiangiogenic effect of Zerumbone, three were computational study and five no full text available. Finally, only 43 articles met the inclusion criteria and were further analysed in this review. The summary of search strategy was depicted in Figure 1.
The number of studies conducted on Zerumbone is strongly correlated to the geographical distribution of Zingiber zerumbet. The highest number of studies conducted on Zerumbone was recorded from Malaysia (15 studies), followed by India (7 studies), China (7 studies), USA (5 studies), Korea (4 studies), Japan (1 study) and Taiwan and Saudi (each of them 2 studies)
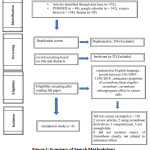 |
Figure 1: Summary of Search Methodology. |
Classification of Included Articles Describing Anticancer and Anti-Angiogenic Activities of Zerumbone
The total number of articles included in this systematic review is 43. The articles were classified according to the models of study applied; animal in vivo, in vitro and ex vivo. Two in vivo studies using animal models, 34 in vitro studies, and 7 studies combining in vitro and in vivo models. Two of the articles had combined ex vivo, in vivo and in vitro studies reported on the anti-angiogenic effect of Zerumbone.
In Vitro Studies with Anti-Cancer and Angiogenetic Effect
Thirty-four in vitro studies have been selected for this review. Majority of the papers are from tropical countries that are rich in ginger plant. Malaysia was the highest country that published articles on this compound.
As summarized in Table I, all studies have demonstrated significant anticancer effect of Zerumbone on wide variety of cell lines. MCF-7 (human breast cancer) is the commonest cell line used (6 articles) followed by Hela (human cervical cancer) (5 articles), HCT116 (human colon cancer cell line) and PC3 (human prostate cancer cell line) which accounted for 4 studies each.
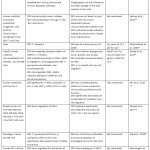 |
Table 1: Summary of in Vitro Anticancer and Antiangiogenic Effects of Zerumbone. |
Discussion
Cell Viability
Cytotoxic evaluation of Zerumbone was carried out in different cell lines. The IC50 (the concentration needed to decrease cell viability by 50% after incubation for 72 h) value was reported in 25 articles. Interestingly, the included studies showed that Zerumbone can affect all tested cell lines and reduced the cell viability of cancer cells in a dose dependent fashion. 8 It is worth to mention that the highest concentration of Zerumbone associated with the lowest cell proliferation in all tested cell lines. The IC50 values was ranged between 3.5 to 46 μM. Zerumbone shown to be less toxic on normal cell line. 8 Range of IC50 value in the selected articles was illustrated in Figure 2.
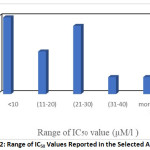 |
Figure 2: Range of IC50 Values Reported in the Selected Articles. |
The antiproliferative activities of Zerumbone showed variable results on a different types of cell lines. The lowest IC50 value was reported by Rajan et al. 28 on CML-K562 cells (chronic myelogenous leukemia cells) which was 3.5 μM, thus suggesting that Zerumbone might be significantly toxic on chronic myeloid leukemia.
Furthermore, Al-Zubairi 8 found that Zerumbone was effective on WEHI-3B (murine monomyelocytic leukemia), 3T3 (mouse fibroblast) and Hela (Human cervical cancer) cell lines with 5, 6 and 6.4 μg/mL as respective IC50 values, followed by CEMss cell line (T4-Lymphoplastoid) with an IC50 value of 12 μg/mL. This proposes the possibility of using Zerumbone in the treatment of hematological cancers and the cervical cancer.
Induction of Apoptosis and the Morphological Changes Induced by Zerumbone
Apoptosis is a biological phenomenon which is defined as programmed cell death, the findings from this indicated the role of Zerumbone in triggering apoptosis. Twenty-seven studies involved reported on apoptosis as an anti-cancer effect of Zerumbone. Abdel Wahab et al. 4 showed morphological changes of cell apoptosis after Zerumbone treatment by using scanning electron microscopy (SEM). In addition, Zerumbone caused significant elevation in the cellular level of caspase-3 in treated cancer cells, as well as an increase in the number of TUNEL-positive stain, which are also characteristics of apoptosis. Similarly, Samad et al. 5 and Hosseinpour et al. 31 described Zerumbone-treated cell morphological changes in cancer cell lines which further suggested that Zerumbone induced apoptosis. Those morphological changes included membrane blebbing and hole formation, cell shrinkage, increase in surface irregularities and apoptotic body formation.
On the other hand, Rajan et al. 28 confirmed apoptosis in Zerumbone treated-CML (chronic myeloid leukemia) cells by demonstrating elevation of signaling molecules which have been responsible on the induction of apoptosis (pro-caspase-3, -9 activation) and poly ADP-ribose polymerase (PARP) cleavage in western blot analysis, the latter is a hallmark of DNA damage.
Apoptosis Signaling Pathways of Zerumbone
Despite of a great number of studies intended to understand signaling pathways of Zerumbone on cancer cell, the exact mechanism is still unclear. So far, there were multiple pathways targeted by Zerumbone recorded in the articles. This can explain the capability of Zerumbone in treating different type of human cell cancers.
Several findings reported various signal pathways targeted by Zerumbone. Nine studies demonstrate caspase 3 and caspase 9 activation as the key in Zerumbone induced apoptosis. Furthermore, six studies stated that Zerumbone induced cell cycle arrest in the G2/M phase. Zerumbone also showed potential in upregulating production of ROS (reactive oxygen species), downregulating protein BCl-2 (anti-apoptotic protein) and upregulating of Bax protein (pro-apoptotic). These were explained in 4 studies. Several studies also substantiated that the loss of mitochondrial membrane potential and caspase activation dependent on cytochrome c was frequently observed in cell death induced by Zerumbone. The possible apoptosis signalling pathways of Zerumbone is illustrated in Figure 3.
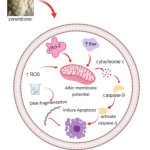 |
Figure 3: Apoptosis Signaling Pathways of Zerumbone. |
Numerous biochemical processes have been distinguished as of great consequence in apoptosis and the key mediator for this process is caspases with caspase-3 as a main promoter of apoptosis. Abdel Wahab et al. 4 stated that Zerumbone induced significant elevation of caspase-3 that play crucial role in the initiation of apoptosis. This finding was supported by Samad et al. 5 results, which stated that the stimulation of caspase-3 and-9 showed by Zerumbone treatment in HepG2 cells. This indicated that Zerumbone induced apoptosis caused by the mitochondrial death pathway.
Moreover, sensitization of mitochondria can be driven under certain condition such as oxidative stress and, upshot of reactive oxygen species known activator of MPTP (mitochondrial permeability transition pore). Opening of these pores will lead to mitochondrial depolarization. Mitochondria will not be able to produce ATP anymore, and thus, the cell will die. Sithara et al. 10 reported that Zerumbone exposure will induce generation of reactive oxygen species, decrease the cellular antioxidant status and loss of mitochondrial membrane potential, the mitochondria membrane potential loss was preceded to initiate the intrinsic pathway of apoptosis through stimulate pro caspase 9 and activated caspase 3.
Correspondingly the data from Chan et al. 21 suggested that Zerumbone induces a crosstalk between endoplasmic reticulum (ER) stress which supported by upregulation of GRP-78 ER chaperone protein that induce assembly of newly-synthesized proteins), upregulation of CHOP expression (a transcription factor that regulates apoptosis in response to cellular stress), and rapid raise of intracellular Ca2+ levels together with the active calpain I (a protein belonging to the family of calcium-dependent proteolytic enzyme) formation. Through calpain and caspase-mediated cleavage of autophagy-regulated proteins Zerumbone can induce autophagy and apoptosis at the same time, this may have been explained by autophagic proteins implied to in regulate mitochondria-involved intrinsic apoptosis. Furthermore, Zerumbone suppressed the assembly of tubulin and elevated the expression of MPM-2 (anti-mitotic proteins).
Similarly, Sobhan et al. 32 reported that the activation of reactive oxygen species as well as calpains is the essential early step in cell death induced by Zerumbone. Mitochondrial pathway was the main pathway in Zerumbone induced apoptosis.
Zerumbone induce apoptosis mainly through intrinsic pathway. The fundamental step in this way is of mitochondrial outer membrane permeabilization. This essential event was controlled by Bcl-2 family. Zerumbone implicated in decreasing the expression of anti-apoptotic proteins such as Bcl-2, Bcl-xL proteins, and increasing the expression of proapoptotic Bax protein. Additionally, overexpression of Bcl 2 family proteins such as Bcl 2 and Bcl-XL generally inhibit Bax dependent cell death by Zerumbone. On the other hand, in cells deficit of Bax expression (Bax knockout cells), Zerumbone failed to induce apoptosis or mitochondrial permeabilization.9,31,32,44
In the other hand, Zainal et al. 11 and Zhou et al. 17 demonstrated that Zerumbone had significant cytotoxic effect. Production of the reactive oxygen species (ROS) induced by Zerumbone, stimulated releases of cytochrome c (cyt-c) into the cytoplasm which has ability to downregulate mitochondrial membrane potential (MMP), Zerumbone increased Bax/Bcl-2 ratio, and concomitant with caspase-3 activation.
Yodkeeree et al. 37 reported the role of TRAIL (tumour necrosis factor–related apoptosis-inducing ligand) in apoptosis induced by Zerumbone. It mediated the activation of extracellular signal-regulated kinase 1/2 and p38 mitogen activated protein kinase which results in DR4 production and DR5 induction. This process then enhances the anticancer effects of TRAIL. TRAIL is one of the tumour necrosis factor family, it is postulated to regulate cancer cells death through different receptors, such as DR4 and DR5, which are transmembrane receptors mediating apoptosis. Stimulation of DR4 and DR5 will result in caspase-8 activation, which will lead to caspase-9 and caspase-3 activation.
Zerumbone also induced p53 protein expression. P53 is tumour suppressor gene with a critical role in regulating apoptosis in cancer cells. Despite that, p53 expression has been linked to the expression of DR4 and DR5. DR5 expression induced by Zerumbone was mediated through a p53-independent mechanism. 9,37
Zerumbone also shows anticancer activity in PANC-1 cells (pancreatic cancer cells) via promoting the upregulation of p53 protein expression and increasing the p21 and miR-34 level, ROS production was shown to be elevated by nearly 149% in PANC-1 cells. It was previously established that p53 activation, induced the increase expression of miR-34 and p21. Zerumbone increased miR-34 and p21 in Zerumbone-treated cancer cells, overexpression of miR-34 preceded growth arrest in cancer cells. 35
Zerumbone has the potential to be used as an adjuvant to potentiate immunotherapy of advanced stages of breast cancer, through inhibition of CD1d expression. CD1d is a lipid antigen presenting molecule and is an indicator for recognition by NKT cells. Advanced stage of breast cancer is associated with increased CD1d expression by the cell. Zerumbone reduced CD1d expression by modifying the antigen presentation pathway and thereby inhibiting cell proliferation. 14 Obviously, these findings confirmed that Zerumbone targeting multiple pathways which contribute to its anti-cancer properties.
Cell Cycle Analysis on Zerumbone
The review showed that Zerumbone induced growth inhibition, and showed that the effect of Zerumbone is refereed to changes in the recognized phase of cell cycle. Eight studies mentioned that Zerumbone arrests cancer cells in the G2/M phase of the cell cycle. The cell cycle confines a chain of events leading to division and replication. G2/M phase is a DNA damage checkpoint . It involves an arrest of the cell in G2 just before mitotic entry in response to stress. 46 Cancer cells divide at an uncontrollable way, and usually lose checkpoint controls that regulate replication of these cells. The effect of Zerumbone in arresting the cell cycle at the G2M phase is a proof to its inhibitory effect on cancer cells proliferation.
Contrary result was reported by Jorvig and Chakraborty 24 and Zhou et al. 17 They demonstrated that Zerumbone significantly induced G1 cell cycle (first growth phase or post mitotic gap phase) arrested followed by apoptosis in SiHa cells (human cervical cancer cells) and DU145 cells (human prostate cancer cell line).
Zerumbone Suppresses Cancer Metastasis and Invasion
Four articles including Kang et al.25, Han et al. 30, Sung et al. 39 and Wang et al. 47 reported that Zerumbone reduced ability of cancer cells to invade and metastasize. 4 different mechanism was described including upregulation of E-cadherin ( cell adhesion proteins), downregulation of CXCR4 (chemokine receptor type 4) expression and a decrease in the osteopontin-induced ROCK/LIMK/cofilin signalling pathway. Zerumbone also suppressed the expression of IL-1ß induced the expression of IL-8 and MMP-3 (matrix metalloproteinase-3) in the breast cancer cell line Hs578T.
Cadherins belong to a family of transmembrane glycoproteins among which E-cadherin withstand. This family is responsible for calcium-dependent intercellular adhesion. Low expression of E-cadherin might promote cancer metastasis. Zerumbone significantly increased expression of E-cadherin and by that reduce cancer metastasis. 47
Rac1 (Ras-related C3 botulinum toxin substrate1) protein is member of the Rac subfamily under the Rho family. Rac1 participates in regulation of several cellular processes including tumour cell invasion and migration as well as cell to cell adhesion. It is highly expressed in many tumours. Wang et al. 47 demonstrated that Rac1 protein is down-regulated when cell migration in inhibited by Zerumbone. Rac1 knockdown could strengthen inhibition of cell migration by Zerumbone. Besides that, Rac1 promotes EMT (Epithelial-mesenchymal transition) and its expression level corresponds with tumour metastases and progression. Zerumbone inhibits cancer cell metastasis by inhibiting EMT process.
CXCR4 and Akt play a versatile part in cancer growth. Both, CXCR4 and Akt, have been involved in cell proliferation, migration and invasion. Zainal et al. 11 stated that Zerumbone repressed OSCC cells (oral squamous cell carcinoma) migration and invasion through suppressing CXCR4 protein expression. Suppression of the CXCR4-RhoA pathway was associated with inhibition of the PI3K-mTOR pathway via Akt and S6 protein inactivation.
There was a number of evidences that supported CXCR4’s role in acting as a marker of cancer aggressiveness. Zerumbone inhibited metastases in OSCCs through the suppression of CXCR4 signalling axis via IL-6 and IL-8 inhibition was previously reported. The finding from Zainal et al. 11 was consistent with Sung et al. 39, where Sung mentioned that Zerumbone downregulated CXCR4 protein expression which led to CXCL12 suppression. CXCL12 is strongly chemotactic. It regulates hematopoietic cell migration, and that Zerumbone inhibited breast and pancreatic cell invasion.
Osteopontin (OPN) is a protein responsible for early T-lymphocyte activation. OPN induces invasion of human non-small cell lung cancer cells.25 OPN inactivates cofilin through the Rho-associated kinase and activated LIM kinases (ROCK/LIMK) pathway. Rho-associated kinase and activate LIM kinases is moderated by the FAK/AKT pathway. OPN and OPN-regulated signalling pathways could play a crucial role for successful treatment of cancer. Kang et al. 25 suggested that invasion of A549 cells was induced by osteopontin and that Zerumbone repressed OPN-induced protein expression of ROCK1, the phosphorylation of LIM kinase 1 and 2 (LIMK1/2), and also cofilin. Through that, Zerumbone was able to strongly suppress the invasion of A549 cells (human non-small cell lung cancer cell). Invasive breast cancers display high amounts of MMP-3, whereas non-invasive human breast cancer cells have undetectable amounts of MMP-3. MMP (Matrix metalloproteinases) are proteolytic enzymes which degrades components of protein in the extracellular matrix and basement membranes. MMPs play a crucial role in cancer invasion and metastasis. IL-1β plays a role as a major proinflammatory cytokine in most types of cancer. IL-1β and TNF-α augments the secretion of MMPs, which gives rise to EMT in human breast cancer growth. Zerumbone in this case was showed to be a potential inhibitor in the expression of IL-8 and MMP-3 leading to the suppression of cell migration and invasion in TNBC cells (breast cancer cell line) induced by IL-1β. 30
The Enhancement of Sensitivity to Radiotherapy by Zerumbone
Deorukhkar et al. 22 described that Zerumbone increase radiation-induced apoptosis by inducing G2/M phase cell cycle arrest. Deorukhkar et al also mentioned that Zerumbone has little apoptotic effect by itself, but it significantly stimulates DNA damage induced by radiation, which was apparent by delayed resolution of post-irradiation nuclear cH2AX foci, whereas Zerumbone alone did not induce cH2AX foci formation. Zerumbone-mediated radiosensitization cellular glutathione (GSH) depletion.
Choi et al. 45 also agreed that pre-treatment with Zerumbone before radiation will induce sensitization to radiation, the authors found that Zerumbone induced sensitization by inhibiting HSP27 and apoptotic molecules (cytochrome c and Protein kinase C delta type) affinity to bind.
Genotoxicity Studies of Zerumbone
Zerumbone had no notable effect on normal human cells, the comet assay results by Al-Zubairi 18 showed that high concentration of Zerumbone could produce DNA damage. Furthermore, results from Al-Zubairi et al. 2 found that there was a direct relationship between the Zerumbone dose and the frequencies of micronuclei (MN assay is a widely used to assess in vitro chromosomal damage). This suggests that high concentrations of Zerumbone could be genotoxic, besides to its cytotoxic effect.
In Vivo Cell Proliferation and Angiogenesis Studies on Zerumbone
The mice model was the most common model used to study the antiangiogenesis and anticancer effect of Zerumbone. Five studies used mice model and another 3 studies used rat model as shown in table II. According to Ni, 43 Zerumbone potentially reduced the tumor size and lung metastasis in C57 BL/6 mice. Additionally, Zerumbone was also found to cause apoptosis and autophagy in melanoma cell lines.
Prenatally exposed Female Balb/c mice to diethylstilbestrol were used in one of the selected studies in order to investigate the anticancer properties of Zerumbone. 40 Results from the experiment showed that Zerumbone effectively induces apoptosis in cervical neoplasia in diethylstilboestrol treated female Balb/c mice.
Park et al. 26 demonstrated marked reduction in hemoglobin content and in vascularization matrigel plug assay in Zerumbone-treated mice. These results indicated that Zerumbone inhibited cancer cell proliferation through different mechanisms which might include its registered antiangiogenics effects.
Furthermore, Kim et al. 41 investigated the effectiveness of dietary administration of Zerumbone, where their result showed that mouse colon and lung carcinogenesis was inhibited by Zerumbone via various modulatory mechanisms of apoptosis, anti-inflammation and suppression of expression of NFϰB and HO-I that were implicated in colon and lung carcinogenesis. Moreover, Al-Zubairi 18 who observed polychromatic erythrocytes as well as the ratio of polychromatic to normochromic erythrocytes of male rats using the micronucleus test (MN) of rat bone marrow reported that the acute exposure to Zerumbone resulted in a heighten number of micronuclei in polychromatic erythrocytes (MNPCEs). This suggested that a high dose of Zerumbone may have a genotoxic and cytotoxic effect.
In vivo data from Choi et al. 45 using nude mice after grafting of NCI-H460 (lower resistant to radiotherapy) and NCI-H1299 (higher radio resistant) cells indicated that Zerumbone builds up radio sensitization for both cell lines. Other than that, the ratio of radio sensitization was higher in NCI-H1299 cell-grafted mice.
Zerumbone suppressed DEN/AAF-induced carcinogenesis in rat liver. Taha et al. 6 reported that Zerumbone antiproliferative effect was mediated through reducing oxidative stress and inducing apoptosis via the mitochondrial pathway. Taha et al. reported that serum AFP (alpha-fetoprotein) concentration was significantly lesser in Zerumbone-treated rats. MDA (malondialdehyde) level which is a biomarker of hepatic lipid peroxidation in cancer cells, was considerably reduced in the cancerous rats treated with Zerumbone. Despite treatment with Zerumbone for 11 weeks, no abnormality was detected in the livers of normal rats.
Shanmugam et al. (2015) mentioned that when Zerumbone (I.P.) was administered to athymic nu/nu mice, activation of STAT3 in tumor tissues was supressed and growth of human RCC (renal cell carcinoma) xenograft tumors in athymic nu/nu mice was inhibited without any significant repercussions.
Additionally, Sehrawat et al. 44 stated that in conjunction with the apoptosis induction and suppression of cell proliferation, Zerumbone administration also significantly delayed growth of orthotropic MDA-MB-231 xenografts (an estrogen-independent cell line with mutant p53).
Antiangiogenic Effects of Zerumbone on Cancer Cell Lines
Four selected articles described the antiangiogenic effect of Zerumbone. 5,26,29,39 These results suggested that antiangiogenic effect of Zerumbone had multipath way also. The anti-angiogenic properties of Zerumbone were potentially controlled by CXCR4 expression, VEGF and STAT3. The possible anti- angiogenesis signalling pathways by Zerumbone was illustrated in Figure 4.
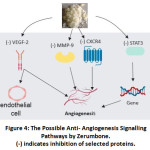 |
Figure 4: The Possible Anti- Angiogenesis Signalling Pathways by Zerumbone. |
Results from Park et al. 26 found that Zerumbone has the potential in decreasing the phosphorylation of vascular endothelial growth factor receptor-2 and fibroblast growth factor receptor-1. These factors help regulate endothelial cell function and angiogenesis. In vivo Matrigel plug assay in Zerumbone treated mice displayed a significant reduction in vascularization counts and haemoglobin content in the plugs. Overall, these results evoke that Zerumbone inhibits different pathways in the angiogenesis process, which might explain its antitumor effects.
According to Sung et al. 39, Zerumbone can reduce CXCR4 expression. This chemokine receptor has been strongly associated to cell proliferation, invasion, angiogenesis, and metastasis in cancer cells. This study suggests that downregulation of CXCR4 expression attributed to down-regulation of NF-ϰB. This leads to CXCL12 inhibition which then inhibits invasion in breast and pancreatic tumour cells as well. Sung also mentioned that Zerumbone inhibits the VEGF inflammatory cytokine which induces CXCR4 expression.
Samad et al. 5 demonstrated that Zerumbone inhibited cell growth and migration of HepG2 cell in a dose‑dependent fashion. This result also showed that Zerumbone decreased expression of angiogenesis molecular effectors such as, the matrix metalloproteinase‑9, vascular endothelial growth factor (VEGF), and VEGF receptor proteins. Additionally, Zerumbone also reduced tube formation by HUVECs through inhibiting new blood vessel and tissue matrix formation.
From other aspect Shanmugam et al. 29 obtained results which established that Zerumbone inhibits STAT3 (signal transducer and activator of transcription) protein which has a crucial role in both angiogenesis and cancer genesis. Shanmugam also suggested that STAT3 activation inhibition by Zerumbone resulted in the expression reduction of the genes involved in proliferation, survival, and angiogenesis which are mediated through protein inducer kinases (c-Src, JAK1, and JAK2 ) inhibition and t phosphatase (SHP-1) induction.
Thus, besides the antiproliferative effects, Zerumbone evidently has anti-angiogenesis effects via inhibition of CXCR4 and MMP, sharing the same affected pathway of antimetastatic effect of Zerumbone. This suggests that Zerumbone inhibits the metastasis of tumours through its antiangiogenic activities.
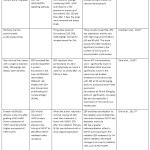 |
Table 2: Summary on the In Vivo Effects of Zerumbone. |
Conclusion
Zerumbone (ZER), present in the subtropical ginger, Zingiber zerumbet has been shown to possess antiproliferative and anti-inflammatory properties. Studies reviewed in this literature proved that Zerumbone can be considered as a potent cancer therapeutic agent. Zerumbone promotes antiproliferative effects through apoptosis induction and cell cycle arrest. Furthermore, this compound exerted ant angiogenesis activities by inhibiting tumor growth and metastasis.
However, this systematic review has number of limitations. The limited data on the toxic dose of Zerumbone is lacking and could be improved by more analyses on the future studies of this compound. Most of the studies on Zerumbone employed in vitro model. In vitro models are generally more susceptible to artefacts due to various variables. The key requirement for in vitro application is validation and quality control. More animal models are required to determine the therapeutic and toxic threshold value of Zerumbone and is essential for translation of drug findings from bench to bedside.
Conflict of Interest
The authors declared there is no potential conflict of interest concerning the research, authorship, and publication of these articles.
Acknowledgments
The authors would like to acknowledge Fundamental Research Grant Scheme, Ministry of Higher Education (MOHE), Malaysia (FRGS/1/2020/STG01/
Funding Sources
Fundamental Research Grant Scheme, Ministry of Higher Education (MOHE), Malaysia (FRGS/1/2020/STG01/
References
- Nikos K and George P. Phosphoinositide 3-Kinase/Akt Signaling and Redox Metabolism in Cancer: Oncol; 2018.
- Al-Zubairi A. S, Abdul A. B, Yousif M, Abdelwahab S. I, Elhassan M. M, Mohan S. In vivo and in vitro genotoxic effects of zerumbone. Caryologia. 2010;63(1):11-17.
CrossRef - Prasannan R, Kalesh K. A, Shanmugam M. K, Nachiyappan A, Ramachandran L, Nguyen A. H, et al. Key cell signaling pathways modulated by zerumbone: role in the prevention and treatment of cancer. Biochem Pharmacol. 2012;84(10):1268-1276.
CrossRef - Abdel Wahab I. S, Abdul A. B, Alzubairi A. S, Mohamed Elhassan M, Mohan S. In vitro ultramorphological assessment of apoptosis induced by zerumbone on (HeLa). Biomed Res Int. 2009;2009:1-10.
CrossRef - Samad N. A, Abdul A. B, Abdullah R, Ibrahim T. A. T, Rahman H, Keong Y. S. Zerumbone (zer) induces apoptosis in Hepg2 cells via mitochondrial pathway. Int J Pharm Pharm Sci. 2015;7(5):599-604.
- Taha M. M. E, Abdul A. B, Abdullah R, Ibrahim T. A. T, Abdelwahab S. I, Mohan S. Potential chemoprevention of diethylnitrosamine-initiated and 2-acetylaminofluorene-promoted hepatocarcinogenesis by zerumbone from the rhizomes of the subtropical ginger (Zingiber zerumbet). Chem Biol Interact. 2010;186(3):295-305.
CrossRef - Rahman H S, Rasedee A, Yeap S. K, Othman H. H, Chartrand M. S, Namvar F, et al. Biomedical properties of a natural dietary plant metabolite, zerumbone, in cancer therapy and chemoprevention trials. Biomed Res Int. 2014;2014:1-20.
CrossRef - Al-Zubairi A. S. Anti-Proliferative activity of zerumbone against tumour cell lines. Online J Biol Sci. 2018;18(2):123-129.
CrossRef - Ma S, Lei Y, Zhang L, Wang J. Effects of zerumbone on proliferation and apoptosis of esophageal cancer cells and on P53 and Bcl-2 expression levels. Oncol Lett. 2018;16(4):4379-4383.
CrossRef - Sithara T, Dhanya B. P, Arun K. B, Sini S, Dan M, Kokkuvayil Vasu R, et al. Zerumbone, a cyclic sesquiterpene from Zingiber zerumbet induces apoptosis, cell cycle arrest, and antimigratory effects in SW480 colorectal cancer cells. J Agric Food Chem. 2018;66(3):602-612.
CrossRef - Zainal N. S, Gan C. P, Lau B. F, San Yee P, Tiong K. H, Rahman Z. A. A, et al. Zerumbone targets the CXCR4-RhoA and PI3K-mTOR signaling axis to reduce motility and proliferation of oral cancer cells. Phytomedicine. 2018;39:33-41.
CrossRef - Hamid A, Rajab N. F, Shen T. S, Nasrom M. N. Cytotoxic and genotoxic effects of zerumbone on WEHI 7.2 wild type murine thymoma cells. J Agric Sci. 2017;9(13):1-13.
CrossRef - Samad N. A, Abdul A. B, Rahman H. S, Rasedee A, Ibrahim T. A. T, Keon Y. S. Zerumbone suppresses angiogenesis in HepG2 cells through inhibition of matrix Metalloproteinase-9, vascular endothelial growth factor, and vascular endothelial growth factor receptor expressions. Pharmacogn Mag. 2017;13(Suppl 4):S731-S736.
- Shyanti R. K, Sehrawat A, Singh S. V, Mishra J. P. N, Singh R. P. Zerumbone modulates CD1d expression and lipid antigen presentation pathway in breast cancer cells. Toxicol In Vitro. 2017;44:74-84.
CrossRef - Thiyam R, Narasu M. L. Zerumbone inhibits proliferation and induces apoptosis, cell cycle arrestin human colon cancer cellline, HCT15. Int J Res Appl Nat Soc Sci. 2017;5(4):49-54.
- Yan H, Ren M. Y, Wang Z. X, Feng S. J, Li S, Cheng Y, et al. Zerumbone inhibits melanoma cell proliferation and migration by altering mitochondrial functions. Oncol Lett. 2017;13(4):2397-2402.
CrossRef - Zhou Y. Z, Tu W. W, Shu C. Z, Gu X. F, Huang Y, Shen X. J, et al. Zerumbone induces G1 cell cycle arrest and apoptosis in cervical carcinoma cells. Int J Clin Exp Med. 2017;10(4):6640-6647.
- Al-Zubairi A. S. Evaluation of cytogenetic and DNA damage effects induced by zerumbone. Asian J Cell Biol. 2016;11(1):13-20.
CrossRef - Jegannathan S. D, Arul S, Dayalan H. Zerumbone, a sesquiterpene, controls proliferation and induces cell cycle arrest in human laryngeal carcinoma cell line Hep-2. Nutr Cancer. 2016;68(5):865-872.
CrossRef - Wang D, Li Y, Cui P, Zhao Q, Tan B-b, Zhang Z-d, et al. Zerumbone induces gastric cancer cells apoptosis: Involving cyclophilin A. Biomed Pharmacother. 2016;83:740-745.
CrossRef - Chan M. L, Liang J. W, Hsu L. C, Chang W. L, Lee S. S, Guh J. H. Zerumbone, a ginger sesquiterpene, induces apoptosis and autophagy in human hormone-refractory prostate cancers through tubulin binding and crosstalk between endoplasmic reticulum stress and mitochondrial insult. Naunyn-Schmiedeberg’s Arch Pharmacol. 2015;388(11):1223-1236.
CrossRef - Deorukhkar A, Ahuja N, Mercado A. L, Diagaradjane P, Raju U, Patel N, et al. Zerumbone increases oxidative stress in a thiol-dependent ROS-independent manner to increase DNA damage and sensitize colorectal cancer cells to radiation. Cancer Med. 2015;4(2):278-292.
CrossRef - Hseu Y. C, Huang Y. C, Korivi M, Wu J. J, Way T. D, Ou T. T, et al. Zerumbone attenuates TGF-β1-mediated epithelial–mesenchymal transition via upregulated E-cadherin expression and downregulated Smad2 signalling pathways in non-small cell lung cancer (A549) cells. J Funct Foods. 2015;18:58-72.
CrossRef - Jorvig J. E, Chakraborty A. Zerumbone inhibits growth of hormone refractory prostate cancer cells by inhibiting JAK2/STAT3 pathway and increases paclitaxel sensitivity. Anticancer Drugs. 2015;26(2):160-166.
CrossRef - Kang C. G, Lee H. J, Kim S. H, Lee E. O. Zerumbone suppresses osteopontin-induced cell invasion through inhibiting the FAK/AKT/ROCK pathway in human non-small cell lung cancer A549 cells. J Nat Prod. 2015;79(1):156-160.
CrossRef - Park J. H, Park G. M, Kim J. K. Zerumbone, sesquiterpene photochemical from ginger, inhibits angiogenesis. Korean J Physiol Pharmacol. 2015;19(4):335-340.
CrossRef - Wang S. D, Wang Z. H, Yan H. Q, Ren M. Y, Gao S. Q, Zhang G. Q. Chemotherapeutic effect of Zerumbone on melanoma cells through mitochondria-mediated pathways. Clin Exp Dermatol. 2016;41(8):858-863.
CrossRef - Rajan I, Jayasree P. R, Kumar P. R. M. Zerumbone induces mitochondria-mediated apoptosis via increased calcium, generation of reactive oxygen species and upregulation of soluble histone H2AX in K562 chronic myelogenous leukemia cells. Tumor Biol. 2015;36(11):8479-8489.
CrossRef - Shanmugam M. K, Rajendran P, Li F, Kim C, Sikka S, Siveen K. S, et al. Abrogation of STAT3 signaling cascade by zerumbone inhibits proliferation and induces apoptosis in renal cell carcinoma xenograft mouse model. Mol Carcinog. 2015;54(10):971-985.
CrossRef - Han J, Bae S. Y, Oh S. J, Lee J, Lee J. H, Lee H, et al. Zerumbone suppresses IL-1β-induced cell migration and invasion by inhibiting IL-8 and MMP-3 expression in human triple-negative breast cancer cells. Phytother Res. 2014;28(11):1654-1660.
CrossRef - Hosseinpour M, Abdul A. B, Rahman H. S, Rasedee A, Yeap S. K, Ahmadi N, et al. Comparison of apoptotic inducing effect of zerumbone and zerumbone-loaded nanostructured lipid carrier on human mammary adenocarcinoma MDA-MB-231 cell line. J Nanomater. 2014;2014:1-10.
CrossRef - Sobhan P. K, Seervi M, Deb L, Varghese S, Soman A, Joseph J, et al. Calpain and reactive oxygen species targets Bax for mitochondrial permeabilisation and caspase activation in zerumbone induced apoptosis. PloS one. 2013;8(4):e59350.
CrossRef - Sun Y, Sheng Q, Cheng Y, Xu Y, Han Y, Wang J, et al. Zerumbone induces apoptosis in human renal cell carcinoma via Gli-1/Bcl-2 pathway. Die Pharmazie-An Int J Pharm Sci. 2013;68(2):141-145.
- Abdel Wahab S. I, Abdul A. B, Zain Z. N. M, Hadi A. H. A. Zerumbone inhibits interleukin-6 and induces apoptosis and cell cycle arrest in ovarian and cervical cancer cells. Int Immunopharmacol. 2012;12(4):594-602.
CrossRef - Zhang S, Liu Q, Liu Y, Qiao H, Liu Y. Zerumbone, a Southeast Asian ginger sesquiterpene, induced apoptosis of pancreatic carcinoma cells through p53 signaling pathway. Evid Based Complement Altern Med. 2012;2012:1-8.
CrossRef - Abdel Wahab S. I, Abdul A. B, Mohan S, Taha M. M. E, Syam S, Ibrahim M. Y, et al. Zerumbone induces apoptosis in T-acute lymphoblastic leukemia cells. Leuk Res. 2011;35(2):268-271.
CrossRef - Yodkeeree S, Sung B, Limtrakul P, Aggarwal B. B. Zerumbone enhances TRAIL-induced apoptosis through the induction of death receptors in human colon cancer cells: Evidence for an essential role of reactive oxygen species. Cancer Res. 2009;69(16):6581-6589.
CrossRef - Abdul A. B. H, Al-Zubairi A. S, Tailan N. D, Wahab S. I. A, Zain Z. N. M, Ruslay S, et al. Anticancer activity of natural compound (Zerumbone) extracted from Zingiber zerumbet in human HeLa cervical cancer cells. Int J Pharmacol. 2008;4(3):160-168.
CrossRef - Sung B, Jhurani S, Ahn K. S, Mastuo Y, Yi T, Guha S, et al. Zerumbone down-regulates chemokine receptor CXCR4 expression leading to inhibition of CXCL12-induced invasion of breast and pancreatic tumor cells. Cancer Res. 2008;68(21):8938-8944.
CrossRef - Abdelwahab S. I, Abdul A. B, Devi N, Taha M. M. E, Al-Zubairi A. S, Mohan S, et al. Regression of cervical intraepithelial neoplasia by zerumbone in female Balb/c mice prenatally exposed to diethylstilboestrol: involvement of mitochondria-regulated apoptosis. Exp Toxicol Pathol. 2010;62(5):461-469.
CrossRef - Kim M, Miyamoto S, Yasui Y, Oyama T, Murakami A, Tanaka T. Zerumbone, a tropical ginger sesquiterpene, inhibits colon and lung carcinogenesis in mice. Int J Cancer. 2009;124(2):264-271.
CrossRef - Samad N. A, Abdul A. B, Abdullah R, Rahman H, Khor K. Z, Chartrand MS. Angiogenic switches play a critical progression in cancer. J Angiotherapy. 2018;1(2):048–055.
CrossRef - Ni Y. In vitro and in vivo studies of pharmacological effects of zerumbone on human malignant melanoma. 2013.
- Sehrawat A, Arlotti J. A, Murakami A, Singh S. V. Zerumbone causes Bax-and Bak-mediated apoptosis in human breast cancer cells and inhibits orthotopic xenograft growth in vivo. Breast Cancer Res Treat. 2012;136(2):429-441.
CrossRef - Choi S. H, Lee Y. J, Seo W. D, Lee H. J, Nam J. W, Lee Y. J, et al. Altered cross-linking of HSP27 by zerumbone as a novel strategy for overcoming HSP27-mediated radioresistance. Int J Radiat Oncol Biol Phys. 2011;79(4):1196-1205.
CrossRef - Yuki K, Junji U, Yusuke C, Nobuyo T, Yoshiko K, Tadaaki Y and Koichi T. Tumor Neovascularization and Developments in Therapeutics: Cancers (Basel). 2019.
- Wang M, Niu J, Gao L, Gao Y, Gao S. Zerumbone inhibits migration in ESCC via promoting Rac1 ubiquitination. Biomed Pharmacother. 2019;109:2447-2455.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






