
Introduction
Dietary oxidized lipids, resulting from frying fats and oils, have been shown to induce negative biological effects.1, 2 Frying oil intensifies the oxidation rate and the unsaturated fatty acids content. During heating and frying the polyunsaturated fatty acids (PUFA) yield more free fatty acids and more polar compounds which alter the fatty acid composition and oil stability, altering the flavour and quality of food.3, 4
Thermal oxidation of unsaturated triglycerides (TAGs)engenders oxidized TAGs, with the formation of decomposition products and polymerized foodstuffs.5 Triglycerides are oxidized and degraded, initially forming oxidized triglycerides3 and upon further oxidation yielding aldehydes and ketones.6 These products negatively impact the quality food,7 and some of the frying compounds are potentially harmful.5
In Argentina, sunflower oil is the most frequent oil used for frying and is consumed in large amounts, fried potatoes being the most consumed deep-fried food.8 Heating this oil produces the highest emissions of total aldehydes5, 9 mainly generated from polyunsaturated triacylglycerols (TAGs) and from other components and reactive intermediates of fried food10 and from the cis-trans isomerization occurring during the deep-fat frying process.11 The intermittent heating and cooling of oils foster increases in mutagenic and carcinogenic products, such as aldehydes.6, 12,13 Furthermore, not only the consumers of fried-food are endangered, those exposed to the repeated cooking with frying oils, especially soy oil or sunflower oil, may also be harmed by the regular exposure to the harmful volatiles compounds generated.14,15 The total polar compounds and distribution of lipids extracted from deep-fried potatoes were like to those obtained from other oils.16
It has been demonstrated that the fried foods intake contributes to the development of chronic diseases such as obesity,17 atherosclerosis,18 liver damage19, 20 and deterioration of bone histomorphometric parameters.21 However, the effects of fried sunflower oil on bone mineral content and bone mechanical competence during growth have not been reported.
In the current study, the negative effect of sunflower-fried food consumption on bone mass and strength during growth was hypothesized. Then, the present study investigated the effect of fried sunflower oil diet on bone mass and bone biomechanical competence of growing rats.
Materials and methods
Animals
Thirty-four male weanling Wistar rats, weighing 46.4 (SD= 5.0) g were provided by the Animal Resources of the Department of Biochemistry, School of Dentistry, University of Buenos Aires, Argentina. The rats were housed in steel individual cages under Laboratory conditions (light-dark 12:12 hours, 21±1ºC and 50–60% humidity). Animals were kept according to the Principles of Laboratory Animal Care established by the National Institute of Health (NIH), USA. Ethics Committee. University of Buenos Aires approved the protocol (UBACyT 20020170100138BA, Nº 004/17).
Diets
The composition of the three diets utilized in the experiment is shown in Table 1. The control diet (C) was a commercial stock diet (Purina chow; Gilardoni SA, Buenos Aires, Argentina). The two experimental diets (SFO and SFOx) differed from the C in fat content and composition as shown in Table 1. Every other day diets were prepared and kept refrigerated. Food intake was measured (Mettler scale PC 4000; accuracy ± 1 mg) and expressed as kcal per 100 g of body weight and per day (kcal/100 g W/day).
C and SFO or SFOx provided 14% and 46% per calories from lipids, respectively. SFO and SFOx were mixed with commercial rat chow at weight ratio of 13% (w/w) of diet. The oils composition of the experimental diets were as follows: (1) SFO composition was: Saturated Fat: 10.1%; Monounsaturated Fat: 27.8%; Polyunsaturated Fat: 61.3%; TFA: 0.7%; (2) SFOx composition: Saturated Fat: 18.09%; Monounsaturated Fat: 33.15%; Polyunsaturated Fat: 46.78%; TFA: 1.66%.
Diets contained calcium carbonate anhydrous at 40.04% (Ca = 143 mg/kg mix), potassium phosphate monobasic at 22.76% (P=44.60 mg/kg mix) and Vitamin D-3 (400 000 IU/g = 250 mg/kg mix) (AIN-93G) 22; 0.6 mg alpha-tocopherol equivalents/g. PUFA were added to the high fat diets as recommended by Valk&Hornstra.23
Frying procedure
Commercial deep-fat fryers with pots (8L) were used for the repeated frozen potato-frying procedure. To assess the oil oxidation, sunflower-oil was heated for 6 hours/day in order to achieve a total of 20, 40 or 60 hours of frying procedure and then, a Testo Equipment, Model 270 at 50ºC was used to analyze the polar compounds (triacylgycerol polymers, dimers, monomers, and oligomers, oxidized triacylglycerols, diacylglycerols and non-esterified fatty acids).
Table 1: Diets composition
| C | SFO | SFOx | |
| Starch | 48 | 30.7 | 30.7 |
| Protein (mix of wheat, corn, fish, soybean and meat flour) | 23 | 14.7 | 14.7 |
| Fat (acid hydrolysis) (mix of fish oil and corn oil) | 7.0 | 4.5 | 4.5 |
| Other sources of fats:Cholesterol (ppm)Linoleic acidLinolenic acidArachidonic acidOmega-3 fatty acidsTotal saturated fatty acidsTotal monounsaturated fatty acids | 2001.30.11<0.011.901.891.98 | 1280.830.071.901.221.27 | 1280.830.071.901.221.27 |
| SFO | 13 | ||
| SFOx | 13 | ||
| FibreMineralsCalciumPhosphorousMagnesiumPotassiumIron (ppm)Copper (ppm)
Zinc (ppm) Sulfur Selenium (ppm) Manganese (ppm) Sodium Chlorine Iodine (ppm) |
6.04.091.200.800.171.10959.0
47 0.14 0.30 25 0.20 0.30 0.60 |
3.82.610.770.500.110.70615.8
28 0.09 0.19 16 0.13 0.19 0.38 |
3.82.610.770.500.110.70615.8
28 0.09 0.19 16 0.13 0.19 0.38 |
| Vitamins mixtureWaterTotal kcal | 1.58.5347.0 | 0.9628.5339.1 | 0.9628.5339.1 |
C: Standard diet; SFO: C diet supplemented with fresh sunflower oil; SFOx: C diet supplemented with thermoxidazed sunflower oil.
At 20, 40 and 60 h, the polar compounds were (n=8; mean ± DS) 20.5±1.02, 29.5±1.77 and 30.5±1.28 g/100g, respectively. Acidity (g oleic acid/100g oil) at 20, 40 and 60 h was 0.110±0.004, 0.160±0.001 and 0.200 ±0.000, respectively. Then, taking into account that frying oils for human consumption must be discarded when polar compounds concentration reaches more than 25% 24-26, the heating time was set at 40 h. Briefly, the frying lasted for 6 h/day at 180ºC; the oil was allowed to cool after each round and it was repeated successively for seven days. Every twelve hours, 500g of potatoes were added to 5 liters of frying oil.
Determination of FA composition of the commercial stock diet and oils
Fatty acid composition of the commercial stock diet, the conventional sunflower oil (SFO) and the thermoxidazed sunflower oil (SFOx) were determined by gas chromatography (GC) technique. Samples preparation were as follows: 100 mg of diet or oil was placed in a tube with a screw cap; 0.35 – 0.40 mL of sodium methoxide solution (0.25 g of metallic sodium in 25 mL of methanol) was added to the tube containing the sample and brought to a bath at 80 °C with shaking for 60 seconds. After 3 minutes, sample was removed from the bath and 0.5 g of the desiccant mixture (calcium chloride – silica gel; 50:50 was added. Then, 3 ml of hexane was added and shacked in vortex for one minute. Sample was centrifuge at 1500 – 1800 rpm and the supernatant containing the methyl ethers was injected into the chromatograph.
Chromatographic conditions were as follows:
Chromatograph Perkin Elmer model Claurus 500, column Supelco SP 2560 (100 m x 0.25 mm x 0.20 μm (a polar biscyanopropyl column specifically designed for detailed separation of geometric positional cis / trans isomers of fatty acid methyl esters – FAMEs-), gas carrier: nitrogen, flame ionization detector (FID, Flame Ionization Detector) at 280 °Con samples derivatized according to Argentine Institute for Standardization and Certification (IRAM) 5650 Part II.27, 28
The fatty acids were identified against patterns SUPELCO F.A.M.E. Mix NHI-C and F.A.M.E. Mix C14-C22; each fatty acid was expressed as the per cent (%) of total fatty acids content.
Experimental design
Rats were randomly assigned to feedings of one of three different diets. The control rats were fed a C diet and the experimental rats received either SFO or SFOx diets. All rats were fed ad libitum throughout the experiment. Food consumption was recorded every other day. At the eight-week experimental period, zoometric and dual-energy x-ray absorptiometry (DXA) analyses were performed under light anaesthesia (5% ketamine hydrochloride; 5 mg/100 g i.p. Holliday–Scott SA. Buenos Aires. Argentina and 2% xylazine hydrochloride; 0.5 mg/100 g i.p. Konig Laboratories. Buenos Aires. Argentina). The animals were euthanized under anaesthesia (0.1 mL of ketamine hydrochloride; 100 mg/mL, Holliday Lab.)/100 g body weight was mixed with 0.02 mL of xylazine; 100 mg/mL, Konig Lab. /100 g body weight) by intramuscular injection. Then, blood samples were collected by cardiac puncture, placed in tubes and the serum was obtained by centrifugation. Serum was stored at –20ºC for biochemical assays. Biochemical determinations and biomechanical assessment were performed in all rats.
Zoometric
Throughout the experimental period, body weights (W, measured in a Mettler PC 4000 scale; accuracy ± 0.001g) and body lengths (L) were registered weakly, after 2 to 4 hours fasting.29
Biochemical Determinations
Blood samples were collected by cardiac puncture at the end of the experimental period. Total serum cholesterol (T-chol; mg/dL of serum), high-density lipoprotein-cholesterol (HDL-chol; mg/dL of serum) and triglycerides (TG; mg/dL of serum) were determined by standardized methods (Roche Diagnostics GmbH, Mannheim, Germany) in a Hitachi 917 autoanalyzer (Hitachi, Tokyo, Japan).
Serum non-HDL cholesterol (mg/dL), a set of atherogenic lipoproteins rich in apo B30, was calculated as the difference between the T-chol and HDL-chol,
due to the loss of sensitivity of this parameter, usually significantly lower than serum HDL-chol levels, in rats.
Serum fatty Acid Composition
Serum fatty acid composition was determined by gas chromatography (GC). Serum fatty acid methyl esters (FAMEs) were determined according to a modified method of Lepage28.
Methanol-n-hexane (2ml; 4:1, vol/vol) was added to 200μl of serum. Then, acetyl chloride (0.2ml) was added and heated for 1h at 100ºC. A solution of K2CO3 (5ml at 6%) was added to the tube, vorterexand centrifuged. The n-hexane top layer containing FAMEs was analyzed by a Chromatograph Perkin Elmer model Claurus 500 and a flame ionization detector (FID, Flame Ionization Detector).
Fatty acids were expressed as the percent (%) of total fatty acids content.
Bone Mass Assessment
Total skeleton bone mineral content (BMC) and bone mineral density (BMD) were assessed in vivo at the end of the experimental period. Under light anesthesia, rats were placed in a total body scanner with software designed specifically for small animals (DPX Alpha 8034, Small Animal Software, Lunar Radiation Corp, Madison, WI), as previously described.31
All rats were scanned using the procedure reported by Zeni et al.,31 The coefficients of variation (CV) evaluated by repeated measurements of five subjects of similar age and gender on five consecutive days for total skeleton BMC and BMD were 3.03 % and 0.89%, respectively. Analysis of spine, femur and lumbar BMC and BMD were performed using a region of interest (ROI) for each segment on the image of the animal on the screen. The CV for spine, femur and lumbar BMD and BMC was 1.79%±0.18. To avoid inter-assay error the same operator performed all DXA measurements.
Biomechanical Testing on Femur
A three-point bending mechanical test (Instron machine, model 4442; Instron Corp) was used to assess right femur biomechanical competence. The bone was positioned horizontally with the anterior side facing down on two transverse supports. Perpendicularly, at the medial aspect of the long axis of the bone, load was applied until fracture. The Instron test machine describes structural bone properties. The resulting load/deformation (W/d) curves measures the bone stiffness and strength. The structural properties are as follows: (i) load at the yielding point (Wy) and marks the initiation of the appearance of microcracks on the periosteal surface of the bone subjected to tension; (ii) bone rigidity (Wydy); and (iii) structural strength (Wf). Geometric or architectural properties are: (i) bone length, (ii) outer vertical and horizontal diameters (V and H, respectively) and (iii) inner vertical and horizontal diameters (v and h, respectively) of the section (Isomet low-speed diamond saw; Buehler).
Measurements were obtained using a stereomicroscope (Stemi DV4; Carl Zeiss Microimaging); accuracy ±0·001 mm. The second moment of inertia of cortical bone area (Ix) was estimated by the following equation: Ix =(π(V3× H −v3× h)/64 mm4). The cross sectional area (CSA) was calculated by a formula (CSA= π/4. (V × H) (mm2)).Femur material properties were estimated from structural and geometrical properties.32 The Young’s modulus of elasticity (E) was calculated using the following formula: E= WyL3/48 dy. Ix (Wy=load at the yielding point, L=distance between supports, dy= maximal elastic deflection, Ix=second moment of inertia of the cross section in relation to the horizontal axis).
Statistical Analysis
All data were analyzed using the statistical software (SPSS v. 20.0 IBM Corp., Chicago, IL. USA) and Graphpad Prism (version 6.0) statistical package (Graphpad).
Data were expressed as mean values with their standard deviations (SD) or standard errors (SE). The data comparisons among the groups was assessed by One-way analysis of variance (ANOVA); statistically significant differences were detected by Student–Newman–Keul’s test or Dunn (not parametric test). Bartlett’s test for homogeneous variances was also done. Normal distribution was verified by Kolmogorov–Smirnov test.P< 0.05 was considered statistically significant (Glantz 1997).
Results
Effects on zoometrics, dietary intake and nutrients
The initial and final body weight and body length of the rats, as well as body weight gain and food consumption of the three groups of rats are shown in Table 2.Allanimals had no significant differences in body weights (P= 0.121) and lengths (P= 0.654) at the beginning of the study. However, body size was significantly affected by the type of dietary fat ingested throughout the experimental period. In fact, body weights and body weight gain per day were significantly lower in the SFOx rats as compared with C and SFO groups (P=0.030) at week 8. The body weights of SFO were similar to the C group (P >0.05).
Moreover, the frying oil consumed altered linear growth. Rats fed SFOx attained the lowest body length (P=0.001), while the SFO and C groups showed similar length growth (P>0.05), over the duration of the experiment.
Rats fed SFO diets containing either fresh oil (SFO) or fried (SFOx) did not show significant differences in energy consumption as compared to C group (P=0.162). However, rats consuming SFO or SFOx diet showed higher fat and monounsaturated fatty acids (MUFA) intake than C rats. SFO and SFOx did not show significant differences. Although SFOx rats consumed higher amounts of SFA and lower of PUFA, including linoleic, linolenic and arachidonic acids than SFO.
Effects on serum lipid profile
The serum lipid profile of the three groups of rats is shown in Figure 1.
The rats fed the SFOx diet showed an altered serum lipid profile as compared to C and SFO groups. SFOx rats showed the highest serum T-chol (P=0.019) and non-HDL-chol (P= 0.008) levels; whereas serum T-chol of SFO and C did not show significant differences between them (P> 0.05).
SFO, SFOx and C groups attained similar serum concentrations of triglycerides and HDL-chol (P = 0.119 and P = 0.208, respectively).
Effects on serum fatty acids
Results of major serum fatty acids found in SFO, SFOx and C groups are summarized in Table 3.
SFOx group presented significantly lower in total PUFA (P = 0.013), 18:2n-6 (P = 0.021), 20:4n-6 (P = 0.040), 22:6n-3 (P = 0.001) profiles than SFO. Total MUFA concentration was similar in the SFOx rats and in the SFO group (P = 0.491), and there was a significant higher profile of total saturated fatty acids in SFOx group (P < 0.014).
Table 2: Zoometrics, dietary intake and nutrients
| CONTROL n=10 |
SFO n=12 |
SFOx n=12 |
P | |
| Initial Body Weight (g) | 46.4±5.0 | 50.5±5.7 | 47.8±6.0 | 0.121 |
| Final Body Weight (g) | 304.9±52.0b | 323.2±23.3b | 278.5±45.2a | 0.030 |
| Body Weight gain (g/day) | 4.61±0.02b | 4.87±0.42b | 4.11±0.66a | 0.001 |
| Initial Body Length (cm) | 13.1±0.6 | 12.9±0.5 | 13.1±0.6 | 0.654 |
| Final Body Length (cm) | 24.4±2.5b | 23.8±0.7b | 22.7±0.8a | 0.001 |
| Energy Intake (kcal/100gW/day) | 45.7±3.1 | 46.4 ± 5.4 | 40.2 ± 10.9 | 0.162 |
| Fat Intake (g/100gW/day) | 0.74±0.1a | 2.3±0.3b | 1.97±0.3b | 0.001 |
| SFA (g/100gW/day) | 0.15±0.01a | 0.24±0.03b | 0.28±0.04c | 0.001 |
| MUFA (g/100gW/day) | 0.30±0.01a | 0.69±0.07b | 0.63±0.06b | 0.001 |
| PUFA (g/100gW/day) | 0.29±0.01a | 1.30±0.02c | 1.13±0.01b | 0.001 |
| Linoleic acid (g/100gW/day) | 0.14±0.01a | 1.20±0.01c | 1.04±0.02b | 0.001 |
| Linolenic acid (g/100gW/day) | 0.01±0.00a | 0.02±0.00c | 0.01±0.00b | 0.001 |
| Arachidonic acid (g/100gW/day) | ———– | 0.004±0.00b | 0.003±0.00a | 0.001 |
Data are means ± SD. Values with different letters are significantly different (P < 0.05) among groups; a being the lowest.
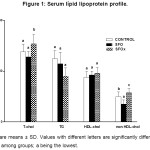 |
Figure 1: Serum lípid lipoprotein profile. Click here to View figure |
Data are means ± SD. Values with different letters are significantly different (P < .05) among groups; a being the lowest.
Table 3: Serum fatty acid (g /100 g of total fatty acids)
| Fatty acid | Control | SFO | SFOx | P | |
| Lauric | 0.82 ±0.01 | 0.40 ±0.42 | 0.59 ±1.00 | 0.583 | |
| Mirístic | 1.80 ±0.37 | 1.32 ±0.59 | 1.83 ±1.05 | 0.483 | |
| Palmitic | 20.49 ±1.33b | 18.32±1.39a | 21.80 ± 2.25b | 0.004 | |
| Palmitoleic | 1.96±0.06b | 1.20±0.25a | 1.35 ±0.27a | 0.001 | |
| Stearic | 14.55±0.37a | 18.61±2.88b | 19.49±2.28b | 0.001 | |
| Oleic | 14.26±0.60 | 14.70±2.19 | 14.65±4.60 | 0.955 | |
| Linoleic | 17.1±0.31a | 20.10±1.89b | 16.40±1.90a | 0.021 | |
| Linolenic | 1.54±0.01b | 0.53±0.14a | 0.48±0.09a | 0.001 | |
| Arachidonic | 12.2±1.77b | 12.34±1.02b | 10.60±1.05a | 0.040 | |
| Eicosapentaenoic | 0.47±0.06a | 0.54±0.10b | 0.59±0.08b | 0.042 | |
| Docosahexaenoic | 1.27±0.19c | 0.99±0.29b | 0.56±0.21a | 0.001 | |
| SFA | 40.0±1.7a | 41.06±5.07a | 46.75±4.66b | 0.014 | |
| MUFA | 19.37±0.89 | 17.82±1.96 | 17.94±4.06 | 0.491 | |
| PUFA | 32.75± 2.16a,b | 34.92±7.04b | 29.02±6.57a | 0.013 | |
| Trans FA | 2.75±0.53a | 3.75±1.09b | 4.19±0.09b | 0.004 | |
Data are means ± SD. Different letters indicate significant differences (P < 0.05) among groups; a being the lowest.
Effects on Bone Mass
The total skeleton body mineral content (BMC), BMC related to body weight (BMC/W), total skeleton body mineral density (BMD); spine BMC/W; spine BMD; femur BMC/W; femur BMD; tibiae BMC/W and tibiae BMD are shown in Table 4.
Rats fed SFOx diet presented significantly diminished total skeleton BMC, BMC/W, femur BMC/W and tibiae BMC/W, as compared with SFO and C groups (P < 0.0001, P < 0.001, P = 0.028 and P< 0.001, respectively).
No significant differences among rats fed SFO and C diets were found for BMC, femur BMC/W and tibiae BMC/W (P >0.05). A decrease in BMC/W was found in the SFO group as compared to C, as shown in Table 4.
There were no differences in BMD, spine BMC/W, spine BMD, femur BMD and tibiae BMD of rats fed either SFO or SFOx, as compared with the C rats (P= 0.329, P= 0.668, P= 0.883, P= 0.599 and P= 0.826, respectively).
Table 4: Bone mass assessment
| CONTROL | SFO | SFOx | P | |
| BMC (g) | 4.846±0.835b | 4.357±0.374b | 2.988±0.516a | 0.000 |
| BMC/W (g/g) | 1.609±0.164c | 1.349±0.070b | 1.192±0.050a | 0.000 |
| BMD (g/mm2) | 0.267±0.007 | 0.257±0.006 | 0.247±0.037 | 0.329 |
| SpineBMC/W (g/g) | 0.620±0.050 | 0.618±0.08 | 0.653±0.100 | 0.668 |
| Spine BMD (g/mm2) | 0.240±0.07 | 0.235±0.08 | 0.217±0.11 | 0.883 |
| Femur BMC/W (g/g) | 0.060±0.007b | 0.063±0.008b | 0.054±0.011a | 0.028 |
| Femur BMD (g/mm2) | 0.296±0.110 | 0.256±0.160 | 0.238±0.006 | 0.599 |
| Tibiae BMC/W(g/g) | 0.358±0.005b | 0.360±0.004b | 0.278±0.011a | 0.000 |
| Tibiae BMD (g/mm2) | 0.245±0.050 | 0.214±0.110 | 0.212±0.130 | 0.826 |
Data are means ± SD. Values with different letters denote significant differences (P< 0.05) between groups; a: being the lowest.
Effects on Bone Biomechanical Competence
The load-bearing capacity and stiffness of the femoral diaphysis are shown in Figure 2.Load-bearing capacity (Wf) and diaphyseal stiffness (Wy/dy) were negatively affected in rats fed sunflower oil either fresh or fried, as compared to the C group (P< 0.001 and P< 0.001; respectively), being significantly lower in rats fed SFOx diet (Figure 2).
The geometrical properties of the femur are shown in Figure 3. The moment of inertia of the fracture section (Ix) and cross-sectional bone area (CSA) were significantly lower in rats fed SFO or SFOx diets than the values found in the C rats (P< 0.001 and P< 0.001; respectively); however, rats fed SFOx diet showed the lowest values (Figure 3).
Young’s modulus of elasticity is shown in Figure 4. There were no significant differences among material properties (P > 0.05). The effects of fried SFO did not influence the material bone quality in terms of yield stress.
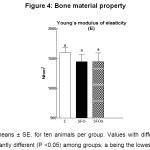 |
Figure 2: Bone structural properties Click here to View figure |
Data are means ± SE. for ten animals per group. Values with different letters are significantly different (P < 0.05) among groups; a being the lowest.
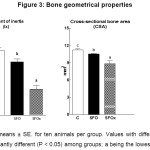 |
Figure 3: Bone geometrical properties Click here to View figure |
Data are means ± SE. for ten animals per group. Values with different letters are significantly different (P < 0.05) among groups; a being the lowest.
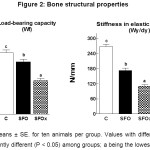 |
Figure 4: Bone material property Click here to View figure |
Data are means ± SE. for ten animals per group. Values with different letters are significantly different (P <0.05) among groups; a being the lowest.
Discussion
This study demonstrated that rats fed a diet containing thermally oxidized sunflower oil fail to gain adequate weight and length and also develop alterations in the serum lipoprotein profiles, bone mass and bone biomechanical competence.
During the frying process, sequences reactions occur in the oil, such as autoxidation, thermal oxidation, polymerization and cyclisation; unsaturated fatty acids are converted to saturated fatty acids and free radicals.19 A large amount of thermoxidazed compounds in the lumen decrease the digestion and absorption33 and the pancreatic lipase activity,34 which negatively affect the hydrolysis of non-oxidized triacylglycerol. The influence of fried-oil consumption on growing rats was also evident by the direct damage by the toxic compounds in the fried-oil associated with histological structural changes. There were also modifications in the microbial profile gut, related to a diminished in total bacterial diversity and density.35-37 In fact, the potential negative effect of diets rich in fried-oil on the intestine could be as a consequence of the lowest bacterial microbiota that may set the stage for a more unstable ecosystem.
The current study indicated that SFOx intake led the rats to achieve the lowest body size, suggesting a negative effect of the components from the fried oil on energy metabolism. These findings are in agreement with Zhou et al.,35 studies, feeding rats with a stock diet plus fried-canola.
Our results suggest that a decrease in food efficiency could also be associated to the damage of essential food nutrients, such as n-6 and n-3 PUFA. Moreover, a diminished in protein, amino acids and fat38 digestive utilization may be due to the reaction of amino acids with primary and secondary products of oxidized lipids. In addition, essential fatty acids deficiency may accompany the ingestion of oxidized fat19,38 and might affect weight gain.39
The rise in total cholesterol and non-HDL-chol serum levels in rats fed SFOx diet may be due to the lower intake of linoleic acid, since PUFA, mainly linoleic acid is recognized to be hypocholesterolemic.38 The hypercholesterolemia observed in this group could be due to the decrease in about 18 % in the serum linoleic acid.
The highest serum cholesterol levels are factors affecting the risk of bone fractures.40, 41 There is also a negative correlation between T-chol and non-HDL-chol serum levels and bone mass.42-44 In human and animals, hypercholesterolemia has been associated with deposition and oxidation of lipids in tissues lead to atherosclerosis45-47and osteopenia.48, 49
On the other hand, the diminished PUFA in SFOx diet was accompanied by an increased total saturated fat. Previous studies showed that rats fed dietary saturated fat impaired bone health in growing rats,50 decreased total skeleton BMD and BMC.51 This was possible due to saturated fatty acids which promote osteoclast survival by inhibiting apoptosis 52with the development of alterations in the serum lipid-lipoprotein profiles.51
It is known that low BMC was considered to increase the risk of developing coronary artery disease.53, 54 The observation of the coexistence of arterial disease and osteoporosis suggests that the same biologically active lipids, lipoproteins and phospholipids involved in atherosclerosis may ultimately affect bone.55 In the present study, SFOx group attained significantly increased in serum T- chol and non-HDL-chol levels and diminutions in femur and tibiae densitometric bone parameters.
Furthermore, the lower bone mass effects in rats fed SFOx was demonstrated by femur cross-sectional-area measurements. Previous studies in vitro have demonstrated that LDL-choloxidation was influenced by the type of dietary oil intake. Moreover, this could have an effect on bone mineralization and bone marrow differentiation.49, 56, 57 The decrease on bone mass in the group fed SFOx was related to the presence of thermoxidazed sunflower rather than the source of oil or the type and concentration of calcium, phosphorous and vitamin D since both SFO and SFOx diets were otherwise similar.
The importance of dietary n-6 and n-3 PUFA is associated, in part, as being precursors of prostaglandins E (PGE). PGE2 derived from n-6 (arachidonic acid, AA)58; PGE3 and Eicosapentaenoic acid (EPA), from n-3 PUFA.59 Moreover, the dual effect of PGE2 on bone metabolism is related to its amount; inhibiting bone formation at high concentrations. In previous studies by us, a higher serum level of AA in rats consuming a rich-linoleic acid diet was observed.50 Then, in the SFO group, a great synthesis of PGE2 -formed by AA60 – rather than PGE3, could be expected.
The lack of decreased bone mass in the SFO group suggests that even though the n-6 content was above the rat requirements, the frying oils and specifically sunflower oil, induce more negative effects on bone than the excess of n-6 PUFA. Moreover, the differences between the SFO and the SFOx dietary intake expose the adverse effects of frying oils on the skeleton of a growing animal rather than the oil per se.
Furthermore, the adverse effect of SFOx on bone mass denoted alterations in bone biomechanical competence. The biomechanical concept of bone quality relates to bone health to bone strength, rather than bone mass. Even though densitometry is unsuitable to assess bone strength and only provides information concerning bone mass, bone quality can be measured by biomechanical competence assays. The results of this study clearly demonstrated adverse effects of ingestion of SFOx diets on the mechanical strength of femoral diaphysis to external loading. Bone stiffness and mass and architectural distribution were more affected in SFOx rats. Bone mechanical quality depends on the mechanical quality of the mineralized tissue (mainly associated with collagen mineralization) and the architectural quality of the structural bone design 32,61
However, bone material quality (E)62 was not modified by fresh sunflower oil or the thermoxidazed oil (SFO or SFOx) diets; this is in congruence with our previous studies in rats fed MUFA-rich diets. 49, 63-65 Since the mechanical quality of the mineralized tissue was not affected, the decreased mechanical competence of the femur of SFO and SFOx rats should be regarded as the result of changes in bone mineral content and spatial distribution of mineralized tissue, rather than bone material properties. Bone with a faster turnover rate (modeling), could reveal potential adverse effects of using thermally oxidized oil in the diet on the skeletal and biomechanical bone properties of a growing animal. 66
Conclusions
The results of this study in growing healthy male rats indicate the prominent role thermoxidazed oil consumption in determining the risk for growth, cardiovascular and bone effects. Consumption of SFOx diet adversely affected body growth, lipid metabolism, bone mass and bone biomechanical competence. The potential adverse effects of thermally oxidized oil in the diet denoted the altered skeletal and biomechanical bone properties of a growing animal in terms of mineralized tissue, rather than bone material properties.
Acknowledgements
We thank Estefania Alsina and Ricardo Orzuza for technical assistance and care of experimental animals. This study was supported by University of Buenos Aires grant UBACyT 20020130100506. Argentina
Conflict of Interest
There was no conflict of interest associated with this study by any of the authors.
References
- Marmesat S., Velasco J., Dobarganes M.C. Quantitative determination of epoxy acids, keto acids and hydroxy acids formed in fats and oils at frying temperatures. Journal of Chromatography. 2008; 1211: 129-134.
CrossRef - Rangel-Zuñiga O.A., Haro C., Tormos C., Perez-Martinez P., Delgado-Lista J., Marin C., Quintana-Navarro G.M., Cerdá C., Sáez G.T., Lopez-Segura F., Lopez-Miranda J., Perez-Jimenez F., Camargo A. Frying oils with high natural or added antioxidants content, which protect against postprandial oxidative stress, also protect against DNA oxidation damage. Eur J Nutr. 2017; 56 (4):1597-1607.
CrossRef - Aniołowska M., Kita A. The effect of frying on glycidyl esters content in palm oil. Food Chem. 2016; 203: 95-103.
CrossRef - Sayyad R. Effects of deep-fat frying process on the oil quality during French fries preparation. J Food Sci Technol. 2007; 4 (8):2224-2229.
CrossRef - Zhang Q., Wan C., Wang C., Chen H, Liu Y, Li S, Lina D, Wu D, Qin W. Evaluation of the non-aldehyde volatile compounds formed during deep-fat frying process. Food Chem. 2018; 243: 151–161.
CrossRef - Zeb A. Chemistry and liquid chromatography methods for the analyses of primary oxidation products of triacylglycerols. Free Radic Res. 2015; 49: 549–564.
CrossRef - Zeb A., Nisar P. Effects of High-Temperature Frying of Spinach Leaves in Sunflower Oil on Carotenoids, Chlorophylls, and Tocopherol Composition. Front Chem. 2017; 5: 19.
CrossRef - Zapata M.E., Rovirosa A., Carmuega E. Changes in the food and beverage consumption pattern in Argentina, 1996-2013 SaludColect. 2016; 12 (4):473-486.
- Peng C.Y., Lan C.H., Lin P.C., Kuo Y.C. Effects of cooking method, cooking oil, and food type on aldehyde emissions in cooking oil fumes. Journal of Hazardous Materials. 2017; 324 (Part B):160-167.
- Zhang Q., Saleh ASM., Chen J., Shen Q. Chemical alterations taken place during deep-fat frying based on certain reaction products: a review. ChemPhys Lipids. 2012; 165: 662-681.
CrossRef - Zhang Q., Wen Q., Lin D., Shen Q., Saleh ASM. The changes in the volatile aldehydes formed during the deep-fat frying process. J Food Sci Technol. 2015; 52 (12):7683-7696.
CrossRef - Dung C., Wu S., Yen G. Genotoxicity and oxidative stress of the mutagenic compounds formed in fumes of heated soybean oil, sunflower oil and lard. ToxicolIn Vitro. 2006; 20:439-447.
CrossRef - Ali RFM, El Anany AM. Recovery of used frying sunflower oil with sugar cane industry waste and hot water. J Food Sci Technol. 2014; 51(11):3002-3013.
CrossRef - Lee T, Gany F. Cooking oil fumes and lung cancer: A review of the literature in the context of the U.S. population. Journal of Immigrant and Minority Health. 2013; 15 (3): 646-652.
CrossRef - Crosa M.J., Skerl V., Cadenazzi M., Olazábal L., Silva R., Suburú G., Torres M. Changes produced in oils during vacuum and traditional frying of potato chips. Food Chemistry. 2014; 146: 603-607.
CrossRef - Marmesat S., Morales A., Velasco J., Dobarganes MC. Influence of fatty acid composition on chemical changes in blends of sunflower oils during thermoxidation and frying. Food Chemistry. 2012; 135: 2333-2339.
CrossRef - Chuang H-C., Huang C-F., Chang Y-C., Lin Y-S., Chao P-M. Gestational ingestion of oxidized frying Oil by C57BL/6 J mice differentially affects the susceptibility of the male and female offspring to diet-induced obesity in adulthood. J Nutr. 2013; 143 (3):267-273.
CrossRef - Xian T.K., Omar N.A., Ying L.W., Hamzah A., Raj S., Jaarin K., Othman F., Hussan F. Reheated palm oil consumption and risk of atherosclerosis: evidence at the ultrastructural level. Evid Based Complement Alternat Med. 2012:828170–828175.
CrossRef - Owu D.U., Osim E.E., Ebong P.E. Serum liver enzymes profile of Wistar rats following chronic consumption of fresh or oxidized palm oil diets. Acta Tropica. 1998; 69: 65-73.
CrossRef - Totani N., Ojiri Y. Thermal deterioration of oil and frying foodstuffs. J Oleo Sci. 2007; 56(10):543-551.
CrossRef - Shuid A.N., Chuan L.H., Mohamed N., Jaarin K., Fong Y.S., Soelaiman IN. Recycled palm oil is better than soy oil in maintaining bone properties in a menopausal syndrome model of ovariectomized rat. Asia Pac J Clin Nutr. 2007; 16 (3):393-402.
- Reeves P., Nielsen F., Fahey G Jr. AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr. 1993; 123:1939-1951.
CrossRef - Valk E.E., Hornstra G. Relationship between vitamin E requirement and polyunsaturated fatty acid intake in man: a review. Int J VitamNutr Res. 2000; 70 (2):31-42.
CrossRef - Firestone D. Regulation of frying fats and oils. In: Perkins EG, Erickson MG, editors. Deep frying: chemistry, nutrition and practical applications. Champaign, IL: AOCS Press; 1996, p. 323-334.
- Hosseini H., Ghorbani M., Meshginfar N., Mahoonak AS. A Review on frying: Procedure, fat, deterioration progress and health hazards. J Am OilChem Soc.2016; 93 (4):445-466.
CrossRef - Zhang Q., Saleh ASM., Shen Q. Monitoring of changes in the composition of soybean oil during deep-fat frying with different food types. J Am OilChem Soc. 2016; 93 (1), 69–81.
CrossRef - Luddy F.E., Barford R.A., Herb S.F., Magidman P. A rapid and quantitative procedure for the preparation of methyl esters of butteroil and other fats. Journal Am Oil Chemists’ Society. 1968; 45, (8), 549–552.
- Lepage G., Roy C.C. Direct transesterification of all classes of lipids in a one-step reaction. J Lipid Res. 1986; 27(1):114-120.
- Vijay K.P., Girish S.J., Parikshit G.B., Mukul J.R. Effect of fasting duration on clinical pathology results in Wistar rats. Vet ClinPathol. 2009; 38:361–366.
CrossRef - Zago V., Lucero D., Macri E.V., Cacciagiu L., Gamba C., Miksztowicz V., Berg G., Wikinski R., Friedman S., Schreier L. Circulating very-low-density lipoprotein characteristics resulting from fatty liver in an insulin resistance rat model. Ann NutrMetab. 2010; 56:198-206.
CrossRef - Zeni S.N., Gregorio S., Gomez A.C., Somoza J., Mautalen C. Olpadronate prevents the bone loss induced by cyclosporine in the rat. Calcif Tissue Int. 2002; 70:48–53.
CrossRef - Turner C.H., Burr D.B. Basic biomechanical measurements of bone: a tutorial. 1993; 14: 595-608.
- Gonzalez-Munoz M.J., Bastida S., Sanchez-Muniz F.J. Short term in vivo digestibility of triglyceride polymers, dimers, and monomers of thermoxidized palm olein used in deep-frying. J Agric Food Chem. 1998; 46: 5188-5193.
CrossRef - Chao P.M., Huang H.L., Liao C.H., Huang S.T., Huang C.J. A high oxidized frying oil content diet is less adipogenic but induces glucose intolerance in rodents. Br J Nutr. 2007; 98 (1): 63-71.
CrossRef - Zhou Z., Wang Y., Jiang Y., Diao Y., Strappe P., Prenzler P., Ayton J., Blanchard C. Deep-fried oil consumption in rats impairs glycerolipid metabolism, gut histology and microbiota structure. Lipids Health Dis. 2016; 15: 86-97.
CrossRef - de La Serre C.B., Ellis C.L., Lee J., Hartman A.L., Rutledge J.C., Raybould H.E. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am J Physiol-Gastr. 2010; 299 (2): G440-448.
CrossRef - Simpson H., Campbell B. Review article: dietary fibre-microbiota interactions. Aliment Pharm Ther. 2015; 42 (2):158-179.
CrossRef - Garrido-Polonio C., García-Linares M.C., García-Arias M.T., López-Varela S., García-Fernández M.C., Terpstra A.H., Sánchez-Muniz F. Thermally oxidised sunflower-seed oil increases liver and serum peroxidation and modifies lipoprotein composition in rats. Br J Nutr 2004; 92 (2):257-265.
CrossRef - López-Varela S., Sánchez-Muniz F.J., Cuesta C. Decreased food efficiency ratio, growth retardation and changes in liver fatty acid composition in rats consuming thermally oxidized and polymerized sunflower oil used for frying. Food Chem Toxicol. 1995; 33 (3):181-189.
CrossRef - Sivas F., Alemdaroğlu E., Elverici E., Kuluğ T., Ozoran K. Serum lipid profile: its relationship with osteoporotic vertebrae fractures and bone mineral density in Turkish postmenopausal women. Rheumatol Int. 2009; 29 (8):885-890.
CrossRef - Peric Kacarevic Z., Snajder D., Maric A., Bijelic N., Cvijanovic O., Domitrovic R., Radic R. High-fat diet induced changes in lumbar vertebra of the male rat offsprings. ActaHistochem. 2016; 118 (7):711-721.
CrossRef - Dennison E.M., Syddall H.E., Aihie Sayer A., Martin H.J., Cooper C. Lipid profile, obesity and bone mineral density: the Hertfordshire Cohort Study. Hertfordshire Cohort Study Group. QJM 2007;100 (5):297-303.
CrossRef - Orozco P. Atherogenic lipid profile and elevated lipoprotein (a) are associated with lower bone mineral density in early postmenopausal overweight women. Eur J Epidemiol. 2004; 19 (12): 1105-1112.
CrossRef - Yamaguchi T., Sugimoto T., Yano S., Yamauchi M., Sowa H., Chen Q., Chihara K. Plasma lipids and osteoporosis in postmenopausal women. Endocr J. 2002; 49 (2):211-217.
CrossRef - Ross R. Atherosclerosis-An inflammatory disease. N Engl J Med. 1999; 340:115-126.
CrossRef - Hmamouchi I., Allali F., Khazzani H., Bennani L., Mansouri L., Ichchou L., Cherkaoui M., Abouqal R., Hajjaj-Hassouni N. Low bone mineral density is related to atherosclerosis in postmenopausal Moroccan women. BMC Public Health. 2009; 9:388-394.
CrossRef - Tamaki J., Iki M., Hirano Y., Sato Y., Kajita E., Kagamimori S., Kagawa Y., Yoneshima H. Low bone mass is associated with carotid atherosclerosis in postmenopausal women: the Japanese Population-based Osteoporosis (JPOS) Cohort Study. Osteoporos Int. 2009; 20:53–60.
CrossRef - Parhami F., Garfinkel A., Demer L.L. Role of lipids in osteoporosis. ArteriosclerThrombVasc Biol. 2000; 20:2346–2348.
CrossRef - Alsina E., Macri E.V., Lifshitz F., Bozzini C., Rodriguez P.N., Boyer P.M., Friedman S.M. Efficacy of phytosterols and fish-oil supplemented high-oleic-sunflower oil rich diets in hypercholesterolemic growing rats. Int J Food SciNutr. 2016; 67(4):441-453.
CrossRef - Macri E.V., Gonzales Chaves M.M., Rodriguez P.N., Mandalunis P., Zeni S., Lifshitz F., Friedman S.M. High-fat diets affect energy and bone metabolism in growing rats. Eur J Nutr. 2012; 51:399-406.
CrossRef - Macri E.V., Lifshitz F., Alsina E., Juiz N., Zago V., Lezón C., Rodriguez P.N., Schreier L., Boyer P.M., Friedman S.M. Monounsaturated fatty acids-rich diets in hypercholesterolemic-growing rats. Int J Food SciNutr. 2015; 66 (4):400-408.
CrossRef - Oh S.R., Sul O.J., Kim Y.Y., Kim H.J., Yu R., Suh J.H., Choi H.S. Saturated fatty acids enhance osteoclast survival. J Lipid Res. 2010; 51(5):892-899.
CrossRef - Barengolts E.I., Berman M., Kukreja S.C., Kouznetsova T., Lin C., Chomka EV. Osteoporosis and coronary atherosclerosis in asymptomatic postmenopausal women. Calcif Tissue Int. 1998; 62:209-213.
CrossRef - Tanko L.B., Bagger Y.Z., Christiansen C. Low bone mineral density in the hip as a marker of advanced atherosclerosis in elderly women. Calcif Tissue Int. 2003; 73:15-20.
CrossRef - Tintut Y., Demer L.L. Effects of bioactive lipids and lipoproteins on bone. Trends Endocrinol Metab. 2014; 25:53-59.
CrossRef - Parhami F., Jackson S.M., Tintut Y., Le V., Balucan J.P., Territo M., Demer LL. Atherogenic diet and minimally oxidized low density lipoprotein inhibit osteogenic and promote adipogenic differentiation of marrow stromal cells. J Bone Miner Res. 1999; 14(12):2067-2078.
CrossRef - Parhami F., Tintut Y., Beamer W.G., Gharavi N., Goodman W., Demer LL. Atherogenic high-fat diet reduces bone mineralization in mice. J Bone Miner Res. 2001; 16(1):182-188.
CrossRef - Raisz L.G. Prostaglandins and bone: physiology and pathophysiology. Osteoarthritis Cartilage 1999; 7:419-421.
CrossRef - Raisz L.G., Alander C.B., Simmons H.A. Effects of prostaglandin E3 and eicosapentaenoic acid on rat bone in organ culture. 1989; 37:615–622.
- Watkins B.A., Li Y., Allen K.G., Hoffmann W.E., Seifert MF. Dietary ratio of (n-6)/(n-3) polyunsaturated fatty acids alters the fatty acid composition of bone compartments and biomarkers of bone formation in rats. J Nutr. 2000; 130 (9):2274-2284.
CrossRef - Ferretti J.L. Biomechanical properties of bone. In: Gennant HK, Guglielmi G, Jergas M, editors. Osteoporosis and bone densitometry. Berlin: Springer Verlag; 1997; p. 143-161.
- Ferretti J.L., Capozza R., Cointry G., Bozzini C., Alippi R.M., Bozzini C.E. Additive effects of dietary protein and energy deficiencies on diaphysis and bone tissue of rat femurs as determined by bending tests. ActaPhysiolPharmacolTherLatinoam. 1999; 41:253-262.
- Boyer P.M., Compagnucci G.E., Olivera M.I., Bozzini C., Roig M.C., Compagnucci C.V., Alippi R.M. Bone status in an animal model of chronic sub-optimal nutrition: a morphometric, densitometric and mechanical study. Br J Nutr. 2005; 93: 663-669.
CrossRef - Pintos P.M., Lezon C.E., Bozzini C., Friedman S.M., Boyer P.M. Operational mechanism modification of bone mechanostat in an animal model of nutritional stress: effect of propranolol. Rev Invest Clin. 2013; 65:39–51.
- Tasat D.R., Lezon C.E., Astort F., Pintos P.M., Macri E.V., Friedman S.M., Boyer P.M. mRNA of cytokines in bone marrow and bone biomarkers in response to propranolol in a nutritional growth retardation model. Pharmacol Rep. 2014; 66:867-873.
CrossRef - Byers S., Moore A.J., Byard R.W., Fazzalari NL. Quantitative histomorphometric analysis of the human growth plate from birth to adolescence. Bone. 2000; 27:495-501.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






