
Introduction
Digestion is the process of breaking down mechanically and enzymatically the food for absorption. 1 This process begins in the mouth, where food is crushed into smaller pieces by the teeth, while both enzymes, such as salivary amylase and lingual lipase, begin chemical digestion. 2 Subsequently, peristaltic contractions of the smooth muscle and mechanical digestion occur in the stomach. Then, further food degradation takes place with the activity of hydrochloric acid and enzymes such as pepsin. 3 Finally, through the action of enzymes such as trypsinogen, pancreatic amylase, pancreatic lipase, chyme trypsinogen, and procarboxypeptidase, among others, chemical digestion in the intestine prepares the food components for absorption by the intestinal cells. 4
Within the process of food digestion, in addition to breaking down nutrients into absorbable elements, during that mechanism, various compounds present in the food matrix of plant-derived products are also released. 5,6 Phenolic compounds are secondary plant metabolites involved in defense against pathogens, protection against herbivores, and antioxidants, among others. 7 Their structure is varied, as they can present one or more hydroxyl groups in their composition and an aromatic ring. 8 Phenolic compounds are classified as simple and polyphenolic, the simple ones have an aromatic ring and an alkyl, aryl, alkenyl, alkoxy, hydroxy or amino group. Polyphenolics, on the other hand have more than one phenolic unit and generally have C15. 9 These compounds are free, bound to organelles, conjugated to macromolecules, confined to cell walls, and trapped in matrices. 10 High bioaccessibility depends on the simple structure of some of the compounds with C6, C6-C1 and others. 11,12 Small phenolic compounds can be absorbed directly in the intestine, whereas complex compounds cannot. 13 Several factors can influence the bioaccessibility of phenolic compounds, the chemical state of the food matrix, interactions or the presence of cofactors or suppressors. It is estimated that 48% of all phenolic compounds are absorbed in the small intestine, 42% in the large intestine and only 10% are not digested. 14 Therefore, the release of these compounds after digestion is called bioaccessibility. Those compounds released from the food matrix that is subsequently absorbed and reach intact or metabolized target tissues are called bioavailable. Likewise, compounds that produce biological effects in the body represent bioactivity. 15
Among the observed benefits of phenolic compounds are the antioxidant activity by neutralizing free radicals resulting from metabolic processes 16, anti-inflammatory effects by reducing the synthesis of proinflammatory cytokines 17,18, as well as cardiovascular 19,20, neurological 21,22, and anticancer effects. 23,24
Due to these health benefits, new foods are developed on whole grains 25,26, fruit and vegetable-based beverages 27,28, fortified dairy products 29,30 and functional foods 31,32, have been developed. However, due to the current conditions on food production and climate change, new sources for food development, which also present beneficial health effects, are being sought. 33-35
Among the new sources of phenolic compounds considered to obtain food responsibly are the use of cereals (sorghum, millet), with low water requirements and resistance to climate change 36, the use of by-products 37,38, as well as the search for new plant resources such as algae and edible halophytes. 39,40
Among the plants, halophytes can develop salt glands, have compatible solute accumulation, specialized root systems, specialized metabolism, and high tolerance to osmotic stress, which allows them to grow in soils with high salt concentrations such as mangroves and coastal areas. 41,42 Due to these stress conditions during their growth, they present considerable concentrations of phenolic compounds 43, exploitable for the development of potentially healthy foods, which in turn would help counteract the effects of climate change (remediation of saline soils) and provide new crops for marginal areas.
Therefore, to know the potential beneficial health effects of halophytes, it is proposed to determine the bioaccessibility of phenolic compounds of S. edulis and S. esteroa, which would bring us closer to understanding the release processes of these compounds during digestion processes.
Materials and Methods
Study area and experimental installation
We carried out this study at the Functional Food Laboratory of the Research and Postgraduate in Food Department (DIPA) of the University of Sonora (UNISON), Campus Centro, Hermosillo, Sonora, northwest Mexico.
Obtaining flour samples
Twenty-five plants of S. edulis and the same number of S. esteroa were collected in Xochimilco, southwest Mexico City, and La Paz, Baja California Sur, Mexico, in December 2021, respectively. We collected plant samples by apical cutting of stems and leaves using pruning shears, then cleaning them with distilled water and subsequently dried and stored in 26.8 x 27.27 cm Ziploc® plastic bags for freezing. To obtain freeze-dried plants, they were frozen in a Thermo Scientific Revco Value Series freezer at -70 °C for 1-h and freeze-dried in a LABCONCO® FreeZone® Freeze Dry System freeze dryer at -50 °C for three days. Next, through a milling and pulverization process, powder with a particle size < 0.45 mm were obtained (Figure 1).
 |
Figure 1: Powders obtained from freeze-dried biomass from S. edulis (A), and S. esteroa (B). |
In vitro simulation gastrointestinal digestion assay
We performed the in vitro simulation gastrointestinal digestion assay following the procedure described Salazar-López et al. (2018) 44, processing the samples of both species, (1) S. edulis and (2) S. esteroa for digestion. The assay consisted of three digestion phases: 1. Oral, 2. Gastric, and 3. Intestinal (Figure 2), following the ethical guidelines of the Council for International Organizations of Medical Sciences. 45 Descriptions of procedures for each phase of digestion are below.
Oral phase
Three asymptomatic fasting volunteers chewed for 15 s 1 g each of S. edulis and S. esteroa meal. Subsequently, they placed each chewed sample in 50 mL Falcon tubes and rinsed their mouths twice with 5 mL of distilled water for 30 s each and expelled the liquid into the corresponding Falcon tubes. Next, they were homogenized and stored on ice for approximately 2 to 3 min.
Gastric phase
For gastric digestion, 5 mL of hydrochloric acid-potassium chloride (HCl-KCl) buffer (2 M, pH 1.5) was added. The pH was adjusted to 1.5 with hydrochloric acid (HCl) (6 M) and sodium hydroxide (NaOH) (2 M) on a HANNA 211 digital pH meter. 667 µL of pepsin solution (300 mg/mL) in HCl-KCl buffer was added. This was then incubated in a shaking water bath Precision Scientific, Winchester, VA, USA, 66800, at 37 °C and 100 rpm for 1 h.
Intestinal phase
For intestinal digestion, 9 mL of saline phosphate buffer (1 M, pH 7.5) was added. The pH was adjusted to 7.5 with hydrochloric acid (HCl) (6 M) and sodium hydroxide (NaOH) (2 M), using the pH meter mentioned above. Then, 1 mL of pancreatin solution (17 mg/mL) in saline phosphate buffer and 80 mg of bile salts were added. Subsequently, it was incubated in the shaking water bath at 37 °C and 100 rpm for 6 h.
 |
Figure 2: General diagram of the in vitro simulation gastrointestinal digestion of S. edulis and S. esteroa. |
Samples obtained in the three digestion steps were centrifuged in a Sigma 3-16KL centrifuge at 4°C and 6000 rpm for 10 min. The supernatants recovered by filtration were frozen in an ARCTIKO ULUF 450 ULUF ultrafreezer at -80°C for 1 h and lyophilized in a LABCONCO lyophilizer at -52°C for 7 days. The lyophilized digests were dissolved in 3 mL of 50% methanol (MeOH). They were then filtered using cellulose acetate membrane filters VWRÒ International North America (0.25 mm, 0.45 μm) and stored at -20 °C, until further analysis.
Determination of total phenols compounds
The content of total phenols compounds of S. edulis and S. esteroa digests was quantified spectrophotometrically by the colorimetric method using the Folin-Ciocalteu reagent (FCR), described by Salazar-López et al. (2016). 46 Briefly, using a NuncTM Edge multiwell plate, 30 µL of each digest was mixed with 150 µL of Folin’s reagent (1:9) and 120 µL of sodium carbonate (Na2CO3) (0.075 mg/mL). The mixture was homogenized and incubated in the dark for 1-h, and absorbance was measured at 765 nm using a FluoStar Omega BMG Labtech Inc. microplate reader spectrophotometer, Ortenberg, Germany. The results were expressed as mg gallic acid equivalents per gram of dry weight sample (mg GAE/g, dw), using a curve as a reference standard.
Determination of total flavonoids compounds
The content of total flavonoid compounds of S. edulis and S. esteroa digests was quantified spectrophotometrically by the colorimetric method using the aluminum trichloride, by Valenzuela-González et al. (2022). 47 We mixed 30 µL of each digest with 9 µL of sodium nitrite (NaNO2) (0.05 mg/mL) and 120 µL of distilled water, using a NuncTM Edge multiwell plate, allowing it to stand for 5 min. Next, 9 µL of aluminum trichloride (AlCl3) (0.1 mg/mL) was added and allowed to stand for another 5 min, adding finally 60 µL of sodium hydroxide (NaOH) (0.04 mg/mL) and 72 µL of distilled water. We stirred the mixture and measured the absorbance at 415 nm using a FluoStar Omega BMG Labtech Inc. microplate reader spectrophotometer, Ortenberg, Germany. The results were expressed as mg quercetin equivalents per gram of dry weight sample (mg QE/g, dw), using a curve as a reference standard.
Determination of Antioxidant Capacity
DPPH ´2,2-Diphenyl-1-picrylhydrazyl´ Assay
We performed a spectrophotometric assay of antioxidant capacity by DPPH (2,2-Diphenyl-1-picrylhydrazyl) of S. edulis and S. esteroa digests, described by Ruiz-Hernández et al. (2021). 25 Subsequently, the working solution of the radical was prepared immediately before use by mixing 2.5 mg of DPPH with 100 mL of methanol (MeOH), the solution obtained in an intense purple color, was adjusted to an absorbance of 0.7 ± 0.02 at 515 nm using a FluoStar Omega BMG Labtech Inc. microplate reader spectrophotometer, Ortenberg, Germany. Using a NuncTM Edge multiwell plate, we mixed 20 µL of each digest with 280 µL of reagent (DPPH radical) (0.025 mg/mL). Once we homogenized the mixture and incubated it in the dark for 1-h, we determined the absorbance changes at the same wavelength using the microplate reader mentioned above. The results were expressed as µmol Trolox equivalents per gram of dry weight sample (µmol TE/g, dw), using a curve as a reference standard.
TEAC ´Trolox Equivalent Antioxidant Capacity ´ Assay
On the other hand, we performed a spectrophotometric assay of antioxidant capacity by TEAC (6-hydroxy-2,5,7,8-tetramethyl-chromium-2-carboxylic acid) of S. edulis and S. esteroa digests, described by Salazar-López et al. (2017). 48 First, we prepared an ABTS activating stock solution by mixing 88 µL of a potassium persulfate (K2S2O8) solution (37.8 mg/mL) with 5 mL of an aqueous ABTS solution (19.3 mg/mL), identifying the solution presented as a deep blue color incubated in the dark at room temperature for 16-18 h. Subsequently, we prepared the working radical solution before use by mixing 1.5 mL of the stock solution with 100 mL of ethanol (C2H6O), adjusting it to an absorbance of 0.7 ± 0.02 at 734 nm with the spectrophotometer mentioned above. Using a NuncTM Edge multiwell plate, we mixed 20 µL of each digest with 280 µL of reagent (ABTS radical). Then, we homogenized the mixture and incubated it in the dark for 5 min, determining the absorbance changes at the same wavelength using the microplate reader mentioned above. The results were expressed as µmol Trolox equivalents per gram of dry weight sample (µmol TE/g, dw), using a curve as a reference standard.
Estimation of Bioaccessibility
We estimated the bioaccessibility of S. edulis and S. esteroa by the concentration of bioactive compounds present in each stage of digestion and the concentration of bioactive compounds found in each extract of the undigested samples; these results are presented in Costa-Becheleni, et al. (2024). 49 The results were expressed in terms of percentage bioaccessibility and recovery of antioxidant activity using the following formula:
Where: BA: bioaccessibility, TP: is the total phenols, TFLA: is the total flavonoids, TAC: is the total antioxidant capacity.
Quantification of Total Phenols and Flavonoids by Ultra High-Performance Liquid Chromatography equipped with a Diode Array Detector (UHPLC-DAD)
Quantification of the total phenol and flavonoid compounds in digests of three phases followed the procedure described by Lee et al. (2012) 50, with slight modifications. We used a ultra high-performance liquid chromatography (UHPLC) system from Agilent Technologies 1260, Germany, equipped with a diode array detector (DAD), separating five phenol acids and five flavonoids with a Zorbax Eclipse Plus-C18 reversed-phase column (2.0 mm x 50 mm 1.8 micron), at a temperature of 30 °C. A two-phase mobile binary elution system of (A) water: formic acid (99.9 : 0.1) and (B) acetonitrile: formic acid (99.9 : 0.1) was applied. A linear elution gradient was as follows:
(1) 0-3 min (97-93 % phase A, and 3-7 % phase B); (2) 3-5 min (93-90 % phase A, and 7-10 % phase B); (3) 5-8 min (90-88 % phase A, and 10-12 % phase B); (4) 8-10 min (88-85 % phase A, and 12-15 % phase B); (5) 10-15 min (85-85 % phase A, and 15-15 % phase B); (6) 15-18 min (85-45 % phase A, and 15-55 % phase B); (7) and 18-20 min (45-10 % phase A, and 55-90 % phase B).
The flow rate was 0.4 mL/min, and the injection volume was 5-μL. To identify phenol and flavonoid compounds, we prepared aqueous methanol extracts by comparing retention time and peaks of the UV spectra of the samples with those of pure reference standards. The results were expressed as µg of phenol acids and flavonoids per gram of dry weight sample (µg phenol ac and fla/g, dw) (using reference curves with each of the acids at different concentrations).
Data Collection and Statistical Analysis
For the numerical analysis, we tabulated the data of the registered variables and classified them in spreadsheets using Excel® 365 version 2022. The results were analyzed with a parametric statistical method, using the Kolmogorov-Smirnov normality test 51 and Levene’s test of similarity of variances test 52 to verify the normal distribution of the data, except for the quantification of phenolic and flavonoid compounds determined by UHPLC-DAD, since statistical programs require at least five data to perform statistical tests and, in this case, we only have three data for each variable. We compared the differences between species with the t-Student test at a 95 % significance level (p < 0.05), generating the resulting graphs with the free statistical software PAST version 4.10. 53
Results and Discussion
Free total phenols and flavonoids during gastrointestinal simulation
Phenolic compounds can be found in different conformations in the food matrix, during the digestion process these can be released to be absorbed and realize their bioactivity or interact with the rest of the food components and digestion elements. 54
Under digestion conditions, the acid pH of the stomach can cause the breaking of ester, glycosidic, and disulfide bonds, causing the release from the food matrix of phenolic compounds. 55 By the effect of pH, denaturation of proteins and depolymerization of polysaccharides can release those compounds that were trapped or bound to them. 56 Likewise, by enzymatic action, hydrolysis of the food matrix structure can cause a release of the compounds trapped in it. 57 On the other hand, the alkaline pH of the intestine can help dissolve the food matrix, releasing phenolic compounds, as well as the presence of enzymes such as pancreatin, consisting of pancreatic amylase and lipase, trypsin, and chymotrypsin, can further degrade the food matrix, predisposing the release of antioxidant compounds. 58-60
Although digestion releases phenolic compounds, hydrolysis of food matrix components will also cause interactions among them. 54 Phenols and proteins can interact, then forming protein-phenol complexes, where these interactions (non-covalent bonds) can affect the structure of proteins and the availability of phenols for absorption. 61 At the same time, phenolic compounds can cause inhibition of digestive enzymes by binding to the active site, inhibiting their bioactivity, which in turn decreases the release of compounds by reducing the number of enzymes that hydrolyze structures where more phenolic compounds are trapped. 54 On the other hand, free phenolic compounds can interact with those carbohydrates hydrolyzed during digestion, forming hydrogen bonds, where hydroxyl groups of phenols and carbohydrates interact; 62 likewise, hydrophobic interactions can occur where non-polar portions can interact with carbohydrates, minimizing the interaction with water. 63 Also, inclusion complexes may occur, where phenols are inserted into the carbohydrate structures, as commonly occurs with the glucose rings in starch. 64
Figure 3 displays the effects of released phenolic compounds from the food matrix, reflecting their interactions. The above reflects an increase of free total phenols and flavonoids during the “in vitro” digestion process, possibly due to enzymatic effects and the pH adjustment, as discussed above.
The results indicate an increase through the different phases; in this sense, the intestinal bioavailable fraction of phenols was higher for S. esteroa, being 4.84 % higher in phenol content compared to S. edulis (18.57 and 17.67 mg GAE/g), showing high significant differences between species (p < 0.05). In contrast, total flavonoid content was statistically equal during all phases; however, the bioavailable intestinal fraction of total flavonoids was higher for S. esteroa, being 0.05% higher in total flavonoid content compared to S. edulis (42.06 and 42.04 mg QE/g), showing non-significant differences (p > 0.05) between species compared to the other phases.
On the other hand, when comparing the content of free phenols of the intestinal fraction with that of both undigested halophytes, we found that after the digestion process, the content of free phenols and flavonoids of the intestinal fraction is 4.84 and 8.01 times more in S. esteroa and 7.68 and 12.51 times more in S. edulis than the content found in the undigested plants.
 |
Figure 3: (A) Total phenols (mg GAE/g, dw: dry weight), and (B) total flavonoids (mg QE/g, dw: dry weight) of oral, gastric, and intestinal phase digests of S. edulis and S. esteroa during the in vitro gastrointestinal simulation process. |
Bioaccessibility of free total phenols and flavonoids during the assessed gastrointestinal simulation process
The results indicate bioaccessibility percentages in S. esteroa of 355.71 for total phenols and 653.90 for total flavonoids, while in S. edulis, the values were 590.16 and 1012.93 % (total phenols and flavonoids, respectively) (Table 1).
Table 1: Estimation of bioaccessibility (%) for total phenols and total flavonoids of oral, gastric, and intestinal phase digests of S. edulis and S. esteroa.
| Species | Oral Phase | Gastric Phase | Intestinal Phase |
| Bioaccessibility (%) of total phenols (mg GAE/g, dw) | |||
| S. edulis | 104.08 | 226.73 | 590.16 |
| S. esteroa | 60.04 | 138.83 | 355.71 |
| Bioaccessibility (%) of total flavonoids (mg QE/g, dw) | |||
| S. edulis | 122.13 | 395.22 | 1012.93 |
| S. esteroa | 78.99 | 259.99 | 653.90 |
Notes. mg GAE/g, dw (mg of gallic acid equivalents/g of sample, dry weight); mg QE/g, dw (mg of quercetin equivalents/g of sample, dry weight); each value is the mean of three replicates.
Evidence suggest that phenols and flavonoids trapped in the food matrix were released in the digestion process. Similar results were found in Hypericum perfoliatum L. after in vitro digestion, where it significantly increases the content of phenols, phenolic acids, and flavonoids. 65 In contrast, different results were observed in quinoa sprouts, where some varieties present lower content of phenols and flavonoids in the intestinal fraction. 66 On the other hand, in Salicornia ramosissima and Sarcocornia fruticosa the highest content of total phenols was found in the gastric phase, which was significantly different. 67
Although the Folin-Ciocalteu technique is widely used to determine the content of total phenols, it is important to consider that this can overestimate or give false positives to the presence of polyphenols, since during extraction it is possible to obtain some reducing compounds such as sugars, 68 and some amino acids 69 that react in the technique. On the other hand, it is important to know that under the aluminum trichloride technique to determine flavonoids can indicate false positives or false negatives, since, being a colorimetric technique, it must be considered that the absorption spectrum of each flavonoid is different, as well as some flavonoids cannot form a complex with aluminum. 70 Therefore, identifying the types of compounds present in the extracts is essential.
Estimation of the total free antioxidant capacity during the gastrointestinal simulation process
The results indicate an increase in antioxidant capacity in all phases; however, the intestinal fractions bioavailable by both colorimetric methods was higher for S. esteroa, being 28.94 % higher in antioxidant capacity content by DPPH (35.03 and 24.89 µmol TE/g) and 23.93 % by TEAC (32.72 and 24.89 µmol TE/g), compared to S. edulis, showing highly significant differences between species (p < 0.05) (Figure 4).
On the other hand, when comparing the antioxidant capacity of the intestinal fraction with that of both undigested halophytes, we found that after the digestion process, the antioxidant capacity of the intestinal fraction increased 1.61 and 1.83 times by DPPH and 1.22 and 1.41 times by TEAC for S. esteroa and S. edulis compared to that found in the undigested plants.
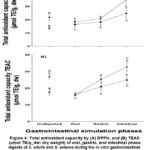 |
Figure 4: Total antioxidant capacity by (A) DPPH, and (B) TEAC (µmol TE/g, dw: dry weight) of oral, gastric, and intestinal phase digests of S. edulis and S. esteroa during the in vitro gastrointestinal simulation process. |
Changes in antioxidant activity during digestion correspond to individual changes in the structure of the phenolic compound. 71,72 In the oral phase, the binding of salivary enzymes to phenols by partial deglycosylation may occur, decreasing the antioxidant activity. 73
The fact that there is no correlation between the content of phenolic and flavonoid compounds and antioxidant activity may be because during chewing and because of salivary enzymes, some of these compounds are released; however, their antioxidant capacity may be much lower than those released during other phases because they are more basic. 74 This may be due to the presence of hydroxyl groups, the substitution of these in the aromatic ring, up to the antioxidant mechanisms that these released compounds may have, such as the of hydrogen atoms transfer (HAT), single electron transfer followed by proton transfer (SET-PT), and sequential protons loss with electron transfer (SPLET). 75 In addition, the synergistic and antagonistic effect of some of these compounds is known, which can positively or negatively influence antioxidant activity. 76
Estimation of antioxidant activity recovery during the gastrointestinal simulation process
Antioxidants such as phenols are molecules involved in free radical scavenging, participating in the inhibition of peroxides and singlet oxygen, and various oxygen reactive species, as they function as chelators of metal ions and scavenge oxidation products. 77
There are different classifications, including primary and secondary, where primary antioxidants inhibit chain oxidation by acting as free radical acceptors or hydrogen donors, which compete with the propagation of lipid oxidation, producing stable products. 78 On the other hand, secondary antioxidants prevent or retard oxidation by suppressing oxidation promoters such as prooxidative enzymes, metal ions, and singlet oxygen. 79 Therefore, determining the antioxidant recovery during the gastrointestinal simulation process brings us closer to the potential beneficial effect of consuming foods rich in phenolic compounds, which could promote some benefits, such as a decrease in arteriosclerosis, aging, and diabetes, among others. 80
The results of the antioxidant recovery during the digestion process of S. edulis and S. esteroa show that during the different phases of digestion, the antioxidant capacity increases, which could be due to the greater availability of phenolic compounds resulting from the hydrolysis of the food matrix. 81 When comparing the antioxidant recovery of the intestinal phase, we recognized the value of halophytes as potentially functional food; however, S. edulis presents significantly higher antioxidant capacity compared to S. esteroa, which could be due to the presence of more different types of antioxidant compounds than S. esteroa. This difference seems to be caused by the extreme growth and stress conditions prevailing in the habitat of these plants. 82 When comparing both plants, antioxidant recovery in DPPH of 157.15 and 181.37 % and in TEAC of 118.07 and 139.74 % were observed for S. esteroa and S. edulis, respectively (Table 2).
Table 2: Estimation of antioxidant capacity recovery total (%) by DPPH and TEAC of oral, gastric, and intestinal phase digests of S. edulis and S. esteroa.
| Species | Oral Phase | Gastric Phase | Intestinal Phase |
| Antioxidant capacity recovery total (%) by DPPH (µmol TE/g, dw) | |||
| S. edulis | 117.85 | 130.58 | 181.37 |
| S. esteroa | 82.44 | 93.91 | 157.15 |
| Antioxidant capacity recovery total (%) by TEAC (µmol TE/g, dw) | |||
| S. edulis | 89.39 | 116.13 | 139.74 |
| S. esteroa | 57.77 | 91.36 | 118.07 |
Notes. DPPH (2,2-Diphenyl-1-picrylhydrazyl); TEAC (Trolox Equivalent Antioxidant Capacity); µmol TE/g, dw (micromoles of Trolox equivalents/g of sample, dry weight); numbers correspond to the mean of three replicates.
A previous study reported different results for Salicornia ramosissima and Sarcocornia fruticosa, where the gastric phase presented the highest antioxidant recovery during the gastrointestinal simulation process. 67 When identifying the antioxidant potential of any extract rich in phenolic compounds, it is necessary to keep in mind that the observed results may be due to different conditions, including antioxidant synergy, 83 antagonistic effects 84 and the chemical structure 85 of the compounds present. Therefore, by identifying each of the compounds individually, it will be known how these possibly increased or decreased the antioxidant potential of the samples.
Content of free phenols and flavonoids found during the gastrointestinal simulation process
After knowing the content of free phenols and total free flavonoids, as well as the antioxidant recovery during the gastrointestinal simulation, by using liquid chromatography, phenolic acids such as ferulic, gallic, p-Coumaric, protocatechuic and synaptic, as well as flavonoids such as catechin, myricetin, naringenin, naringin and ruthin were identified according to the retention times and spectrum of each compound compared with different standards. These compounds were also identified in undigested samples (data not shown).
When comparing the phenolic compounds found in S. edulis and S. esteroa with the content of total phenols and flavonoids, we could attribute that much of its content is given by the identified compounds, however, other compounds that could not be identified could positively influence the antioxidant capacity of these plants, this is because compounds such as caffeoylquinic acid, hexosyl rhamnetin pentoside and dihydroquercetin among others, have been abundantly identified in different halophytes. 67 When observing the content of the bioactive compounds identified, it was found that some phenolic acids and flavonoids increase and decrease through the different gastrointestinal phases, which demonstrates the release and interactions that these compounds have with the food matrix. When comparing what happens with each compound in the intestinal phase, an important phase since it is where compounds would be available for absorption by the intestine to carry their bioactivity, only some compounds such as gallic acid, p-Coumaric acid, catechin, myricetin and ruthin in S. edulis, p-Coumaric, protocatechuic, synaptic, catechin, myricetin, naringenin and ruthin in S. esteroa presented the highest concentration significantly in the intestinal phase compared to the oral and gastric phases.
However, both phenolic acids and flavonoids from the intestinal phase in both plants, the concentration found bioaccessible compared to the concentration of undigested plants is varied according to the different types of compounds. Where most of the compounds except p-Coumaric, catechin and naringin decreased after digestion in S. edulis. Compared to S. esteroa, p-Coumaric acid and ruthin were the only compounds that did not decrease after simulation. On the other hand, when comparing the content of bioactive compounds of halophytes before and after digestion, as observed, most of the compounds presented low percentages of bioaccessibility, however, p-Coumaric acid, catechin and naringin in S. edulis presented very good percentages of bioaccessibility. This reflects the dynamic effect that digestion has on the food matrix and the content of bioactive compounds. The results are shown in Tables 3 and 4.
Table 3: Identification for phenols acids from oral, gastric, and intestinal phase digests of S. edulis and S. esteroa by UHPLC-DAD.
| Species | Compounds (µg/g, dw) | Oral | Gastric | Intestinal |
Bioaccessibility (%) |
|
S. edulis
|
Ferulic | 603.10±82.93 b A | 157.57±5.00 ns C | 178.71±6.37 b B ↓ | 3.99 |
| Gallic | 1967.97±37.66 a C | 3395.58±273.33 a A | 3099.94±213.03 a B ↓ | 91.49 | |
| p-Coumaric | 9.70±0.05 b C | 35.98±0.60 ns B | 428.88±33.71a A ↑ | 488.08 | |
| Protocatechuic | 1.04±0.11 ns C | 2.02±0.18 a B | 9.65±0.67 ns A ↓ | 5.04 | |
| Synaptic | 66.93±7.83 a C | 75.77±2.39 ns A | 75.38±4.51 ns B ↓ | 73.07 | |
|
S. esteroa |
Ferulic | 1114.64±84.00 a A | 118.32±47.97 ns C | 264.74±2.55 a B ↓ | 10.40 |
| Gallic | 1.83±0.20 b A | 1.17±0.05 b B | 1.14±0.09 b C ↓ | 81.42 | |
| p-Coumaric | 30.57±4.66 a C | 37.27±4.45 ns B | 70.63±3.67 b A ↑ | 114.47 | |
| Protocatechuic | 0.99±0.11 ns C | 1.32±0.06 b B | 8.29±0.74 ns A ↓ | 15.46 | |
| Synaptic | 43.95±7.70 b C | 60.04±8.99 ns B | 78.07±4.72 ns A ↓ | 9.09 |
Notes. µg/g, dw (micrograms/g of sample, dry weight); numbers correspond to the mean ± standard deviation of three replicates; superscripts a and b between columns indicate statistically significant differences of the same compound between species; superscripts A, B and C between lines indicate statistically differences of the same compound across digestion phases (t-Student test, p < 0.05); superscripts ns indicate statistically non-significant differences according to the above comparisons (t-Student test, p > 0.05); the arrows ¯ indicate increase or decrease in the concentration of bioactive compounds compared to undigested sample.
Tabla 4: Identification for flavonoids from oral, gastric, and intestinal phase digests of S. edulis and S. esteroa by UHPLC-DAD.
| Species | Compounds (µg/g, dw) | Oral | Gastric | Intestinal |
Bioaccessibility (%) |
|
S. edulis
|
Catechin | 20.47±2.74 a C | 39.79±1.90 a B | 48.91±3.82 a A ↑ | 764.21 |
| Myricetin | 20.37±1.22 a B | 1.93±0.02 ns C | 25.57±0.28 a A ↓ | 15.60 | |
| Naringenin | 32.99±1.59 ns A | 13.24±1.10 b C | 23.30±1.01 b B ↓ | 28.21 | |
| Naringin | 173.92±1.66 b B | 284.47±39.48 a A | 113.19±8.25 b C ↑ | 252.20 | |
| Ruthin | 5956.39±157.77 a B | 4271.25±641.85 a C | 6911.01±711.06 a A ↓ | 44.58 | |
|
S. esteroa |
Catechin | 11.08±1.97 b B | 7.05±0.13 b C | 13.18±0.55 b A ↓ | 10.67 |
| Myricetin | 2.49±0.04 b B | 1.78±0.10 ns C | 3.02±0.29 b A ↓ | 5.51 | |
| Naringenin | 32.92±3.02 ns C | 64.18±5.95 a A | 63.21±7.38 a B ↓ | 63.24 | |
| Naringin | 395.38±46.40 a A | 142.20±17.41 b C | 208.54±31.99 a B ↓ | 80.52 | |
| Ruthin | 0.74±0.04 b B | 0.29±0.04 b C | 8.49±0.34 b A ↑ | 194.41 |
Notes. µg/g, dw (micrograms/g of sample, dry weight); numbers correspond to the mean ± standard deviation of three replicates; superscripts a and b between columns indicate statistically significant differences of the same compound between species; superscripts A, B and C between lines indicate statistically differences of the same compound across digestion phases (t-Student test, p < 0.01); superscripts ns indicate statistically non-significant differences according to the above comparisons (t-Student test, p > 0.05); the arrows ¯ indicate increase or decrease in the concentration of bioactive compounds compared to an undigested sample.
Other authors reported similar results in barley, observing that through digestion, some compounds such as vinylic, ferulic, and p-coumaric acids, among others, vary in their concentration. Therefore, when calculating the release of some compounds, high and low percentages of bioaccessibility of some of them are shown. 86 Similar results were found in wheat, where after digestion, some compounds vary in concentration; likewise, bioaccessibility percentages above 500 % are presented in some compounds. 87
In contrast, in halophytes such as Sarcorconia fruticosa and Salicornia ramosissima compounds, such as caffeoylquinic acids and gallocatechin identified before digestion were not detected in the intestinal phase. 67
The localization of phenolic compounds in different plants can cause a variety of results during the digestion process since some of them are free, can be conjugated or trapped in more complex elements such as proteins, oligosaccharides, polysaccharides, as well as those trapped could be released, but conjugated again. 10 For example, phenolic compounds tend to be conjugated or trapped between proteins and polysaccharides 88,89, whereas those compounds such as ferulic acid tend to be mostly bound to the arabinose chains of plant cell walls 90, while gallic acid is abundantly free in plant cells. 91
Evaluating the effect of digestion on the content of phenols and flavonoids in halophytes such as S. edulis and S. esteroa helps to understand the behavior of these compounds during the gastrointestinal tract, as well as to elucidate the potential beneficial effect that these compounds could have by their consumption. However, even if good bioaccessibility is present, bioactivity in target tissues will depend on bioavailability, as well as the transformations that occur to the bioactive compounds in the liver for being transported in the bloodstream, which could modify the antioxidant potential and beneficial effects. 92,93 With the above considerations, it is possible to use S. edulis and S. esteroa halophytes as potential beneficial foods to counteract some chronic degenerative- diseases, as some bioaccessible compounds in these plants have anticancer properties 94, antidiabetic 95 and other effects. In this context, promoting the production of this kind of plant biomass could help to remediate saline soils, enhancing food production and helping to counteract food insecurity problems. 96
Conclusions
The content of phenols, flavonoids, and antioxidant recovery of halophytes increased during digestion. However, S. esteroa presented significantly higher contents of free total phenols and flavonoids in the intestinal fraction than S. edulis. Meanwhile, S. edulis has higher antioxidant recovery when compared to S. esteroa in the intestinal fraction. Very high percentages of bioaccessibility and antioxidant recovery (> 100%) were present in both halophytes. Gallic acid was the compound with the highest concentration in S. edulis in the intestinal fraction; however, p-Coumaric acid presented the highest percentage of bioaccessibility. In S. esteroa, ferulic acid presented the highest concentration in the intestinal phase. According to these results, we observed considerable bioaccessibility in this plant. Ruthin in S. edulis presented high concentrations in the intestine, while catechin and naringin presented the best bioaccessibility. In S. esteroa, naringin is the most abundant compound, while ruthin presented percentages above 100% bioaccessibility.
The halophytes S. edulis and S. esteroa reflect a high potential as a health food, which could help to counteract chronic degenerative diseases related to obesity and help to solve marginal and saline soils, as well as a potential crop to counteract the problems of food security and climate change.
Acknowledgement
The first author would like to thank the Agricultural Engineer and producer of romeritos in the Chinampa area Prospero Rosales for his support for the field trip to his S. edulis production site. To the technician Pedro Luna García of the Experimental Station of the Center for Biological Research of Northwest Mexico S.C., for his support in the local field trip to the site of establishment, and collection of S. esteroa at Balandra beach, La Paz, Baja California Sur, Mexico. To Dr. Miguel Angel Zavala Sánchez, M. en C. Raúl Calleros Flores of the Autonomous Metropolitan University (UAM-Xochimilco) in the Department of Biological Systems. To Dra. Claudia Judith Hernández Guerrero of the Interdisciplinary Center for Marine Sciences (CICIMAR-IPN) in the Microbiology, and Molecular Biology Laboratory for their support, and assistance in the processing and drying of the samples of edulis, and S. esteroa.
Funding Sources
Part of this research was funded by the National Council for the Humanities, Sciences and Technologies of Mexico (CONAHCyT) through the grant supported with the scholarship number 766309 for the first author’s doctoral studies, which facilitated her stay in Hermosillo, Sonora, Mexico. This project and the APC were funded by Center for Biological Research of Northwest México (CIBNOR), grant number ‘10025-PAZA’.
Conflict of Interest
All authors declare no conflict of interest. The authors have read and approved the final version of the manuscript.
Authors’ Contribution
Each author has significantly and directly contributed intellectually to the manuscript. Author Contributions: FRCB, ETD, RMRS proposed and designed the experiments; FRCB, AARH, RMRS, NJSL performed the lab procedures and methods; FRCB, FAN organized and assisted in field sampling; FRCB, FAN executed the biomass processing; LABS, RMRS helped in interpreting the biochemical and nutritional data; FRCB, ETD, AARH wrote the first version of manuscript. FRCB, ETD, LABS, RMRS reviewed the overall quality of the manuscript.
Data Availability Statement
The manuscript incorporates all datasets produced or examined throughout this research study.
Ethics Statement
The document accurately and thoroughly presents the authors original research and analysis.
Informed Consent Statement
This research did not involve human participants, animal, subjects, or any material that requires ethical approval
References
- Patricia, J. J., Dhamoon, A. S. Physiology, digestion. Eur PMC. In: StatPearls Publishing, Treasure Island (FL) 2019; PMID: 31334962. Available from: https://europepmc.org/article/nbk/nbk544242
- Lai, W. Y. W., Chua, J. W. M., Gill, S., Brownlee, I. A. Analysis of the lipolytic activity of whole-saliva and site-specific secretions from the oral cavity of healthy adults. Nutrients. 2019; 11(1):191. DOI: https://doi.org/10.3390/nu11010191
CrossRef - Schubert, M. L. Physiologic, pathophysiologic, and pharmacologic regulation of gastric acid secretion. Curr Opin Gastroenterol. 2017; 33(6):430-438. DOI: 1097/MOG.0000000000000392
CrossRef - Pandiri, A. R. Overview of exocrine pancreatic pathobiology. Toxicol Pathol. 2014; 42(1):207-216. DOI: https://doi.org/10.1177/0192623313509907
CrossRef - Kumar, N., Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol Rep. 2019; 24:e00370. DOI: https://doi.org/10.1016/j.btre.2019.e00370
CrossRef - Angelino, D., Cossu, M., Marti, A., Zanoletti, M., Chiavaroli, L., Brighenti, F., Martini, D. Bioaccessibility and bioavailability of phenolic compounds in bread: A review. Food Funct. 2017; 8(7):2368-2393. DOI: https://doi.org/10.1039/C7FO00574A
CrossRef - Kołton, A., Długosz-Grochowska, O., Wojciechowska, R., Czaja, M. Biosynthesis regulation of folates and phenols in plants. Sci Hortic. 2022; 291:110561. DOI: https://doi.org/10.1016/j.scienta.2021.110561
CrossRef - Kumar, S., Abedin, M. M., Singh, A. K., Das, S. Role of phenolic compounds in plant-defensive mechanisms. In: Plant Phenolics Sustainable Agriculture, Lone, R, et al. Eds., Springer, Singapore. 2020; 1:517-532. DOI: https://doi.org/10.1007/978-981-15-4890-1_22
CrossRef - Al Mamari, H. H. Phenolic compounds: Classification, chemistry, and updated techniques of analysis and synthesis. Phenolic compounds-chemistry, synthesis, diversity, non-conventional industrial, pharmaceutical, and therapeutic applications 2021; 10. Available from: https://www.intechopen.com/chapters/77604
CrossRef - Parada, J., Aguilera, J. M. Food microstructure affects the bioavailability of several nutrients. J Food Sci. 2007; 72(2):R21-R32. DOI: https://doi.org/10.1111/j.1750-3841.2007.00274.x
CrossRef - Vuolo, M. M., Lima, V. S., Junior, M. R. M. Phenolic compounds: Structure, classification, and antioxidant power. In Bioactive Compounds, Woodhead Publishing. 2019; 33-50. DOI: https://doi.org/10.1016/B978-0-12-814774-0.00002-5
CrossRef - Ribas-Agustí, A., Martín-Belloso, O., Soliva-Fortuny, R., Elez-Martínez, P. Food processing strategies to enhance phenolic compounds bioaccessibility and bioavailability in plant-based foods. Crit Rev Food Sci Nutr. 2018; 58(15):2531-2548. DOI: https://doi.org/10.1080/10408398.2017.1331200
CrossRef - Lippolis, T., Cofano, M., Caponio, G. R., De Nunzio, V., Notarnicola, M. Bioaccessibility and bioavailability of diet polyphenols and their modulation of gut microbiota. Int J Mol Sci. 2023; 24(4):3813. DOI: https://doi.org/10.3390/ijms24043813
CrossRef - Wojtunik-Kulesza, K., Oniszczuk, A., Oniszczuk, T., Combrzyński, M., Nowakowska, D., Matwijczuk, A. Influence of in vitro digestion on composition, bioaccessibility and antioxidant activity of food polyphenols—A non-systematic review. Nutrients, 2020; 12(5):1401. DOI: https://doi.org/10.3390/nu12051401
CrossRef - Rodrigues, D. B., Marques, M. C., Hacke, A., Loubet Filho, P. S., Cazarin, C. B. B., Mariutti, L. R. B. Trust your gut: Bioavailability and bioaccessibility of dietary compounds. Curr Res Food Sci. 2022; 5:228-233. DOI: https://doi.org/10.1016/j.crfs.2022.01.002
CrossRef - Eseberri, I., Trepiana, J., Léniz, A., Gómez-García, I., Carr-Ugarte, H., González, M., Portillo, M. P. Variability in the beneficial effects of phenolic compounds: A review. Nutrients. 2022; 14(9):1925. DOI: https://doi.org/10.3390/nu14091925
CrossRef - Limmongkon, A., Nopprang, P., Chaikeandee, P., Somboon, T., Wongshaya, P., Pilaisangsuree, V. LC-MS/MS profiles and interrelationships between the anti-inflammatory activity, total phenolic content, and antioxidant potential of Kalasin 2 cultivar peanut sprout crude extract. Food Chem. 2018; 239:569-578. DOI: https://doi.org/10.1016/j.foodchem.2017.06.162
CrossRef - Ford, C. T., Richardson, S., McArdle, F., Lotito, S. B., Crozier, A., McArdle, A., Jackson, M. J. Identification of (poly) phenol treatments that modulate the release of pro-inflammatory cytokines by human lymphocytes. B J Nutr. 2016; 115(10):1699-1710. DOI: http://dx.doi.org/10.1017/S0007114516000805
CrossRef - Ali, S. S., Ahmad, W. A. N. W., Budin, S. B., Zainalabidin, S. Implication of dietary phenolic acids on inflammation in cardiovascular disease. Rev Cardiovasc Med. 2020; 21(2):225-240. DOI: https://doi.org/10.31083/j.rcm.2020.02.49
CrossRef - Lutz, M., Fuentes, E., Ávila, F., Alarcón, M., Palomo, I. Roles of phenolic compounds in the reduction of risk factors of cardiovascular diseases. Molecules. 2019; 24(2):366. DOI: https://doi.org/10.3390/molecules24020366
CrossRef - Conde, C., Escribano, B. M., Luque, E., Aguilar-Luque, M., Feijóo, M., Ochoa, J. J., Túnez, I. The protective effect of extra-virgin olive oil in the experimental model of multiple sclerosis in the rat. Nutr Neurosci. 2020; 23(1):37-48. DOI: https://doi.org/10.1080/1028415X.2018.1469281
CrossRef - Qneibi, M., Hanania, M., Jaradat, N., Emwas, N., Radwan, S. Greuter, phytochemical composition, antioxidant, total phenolic content, total flavonoids content and neuroprotective effects. Eur J Integr Med. 2021; 42:101291. DOI: https://doi.org/10.1016/j.eujim.2021.101291
CrossRef - Abotaleb, M., Liskova, A., Kubatka, P., Büsselberg, D. Therapeutic potential of plant phenolic acids in the treatment of cancer. Biomolecules. 2020; 10(2):221. DOI: https://doi.org/10.3390/biom10020221
CrossRef - Anantharaju, P. G., Gowda, P. C., Vimalambike, M., Madhunapantula, S. An overview on the role of dietary phenolics for the treatment of cancers. Nutr J. 2016; 15:1-16. DOI: https://doi.org/10.1186/s12937-016-0217-2
CrossRef - Ruiz-Hernández, A. A., Cárdenas-López, J. L., Cortez-Rocha, M. O., González-Aguilar, G. A., Robles-Sánchez, R. M. Optimization of germination of white sorghum by response surface methodology for preparing porridges with biological potential. CyTA J Food. 2021; 19(1):49-55. DOI: https://doi.org/10.1080/19476337.2020.1853814
CrossRef - Leliana, L., Setyaningsih, W., Palma, M., Supriyadi, S., Santoso, U. Incorporation of Young Coconut (Cocos nucifera) Mesocarp Increases the Antioxidant Activity, Phenolic Compounds and Oxidative Stability of Cookies. Trends Sci. 2024; 21(2):7199-7199. DOI: https://doi.org/10.48048/tis.2024.7199
CrossRef - Rigolon, T. C. B., Borges, L. L. R., Nascimento, A. L. A. A., Fernandes, J. G., Marins, J. C. B., Martins, E., Stringheta, P. C. Study of the stability of hydroelectrolytic sports beverages enriched with phenolic extract from jaboticaba peel or blueberry pulp. Food Hum. 2024; 2:100214. DOI: https://doi.org/10.1016/j.foohum.2023.100214
CrossRef - Pinto, T., Vilela, A. Healthy drinks with lovely colors: Phenolic compounds as constituents of functional beverages. Beverages. 2021; 7(1):12. DOI: https://doi.org/10.3390/beverages7010012
CrossRef - Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J Food Biochem. 2020; 44(9):e13394. DOI: https://doi.org/10.1111/jfbc.13394
CrossRef - Zeb, A. Phenolic Antioxidants in Dairy Products. In: Phenolic Antioxidants in Foods: Chemistry, Biochemistry and Analysis, Springer, Cham. 2021; 281-296. DOI: https://doi.org/10.1007/978-3-030-74768-8_10
CrossRef - Ashaolu, T. J., Adeyeye, S. A. African Functional Foods, and Beverages: A Review. J Culin Sci Technol. 2024: 22(1):142-177. DOI: https://doi.org/10.1080/15428052.2022.2034697
CrossRef - Kauser, S., Hussain, A., Ashraf, S., Fatima, G., Javaria, S., Abideen, Z. U., Korma, S. A. Flaxseed (Linum usitatissimum); phytochemistry, pharmacological characteristics and functional food applications. Food Chem Advan. 2024; 4:100573. DOI: https://doi.org/10.1016/j.focha.2023.100573
CrossRef - Lee, C. C., Zeng, M., Luo, K. How does climate change affect food security? Evidence from China. Environ Impact Assess Rev. 2024: 104:107324. DOI: https://doi.org/10.1016/j.eiar.2023.107324
CrossRef - Janni, M., Maestri, E., Gullì, M., Marmiroli, M., Marmiroli, N. Plant responses to climate change, how global warming may impact on food security: a critical review. Front Plant Sci. 2024: 14(2023):1297569. DOI: https://doi.org/10.3389/fpls.2023.1297569
CrossRef - Bedasa, Y., Deksisa, K. Food insecurity in East Africa: An integrated strategy to address climate change impact and violence conflict. J Agric Food Res. 2024: 15:100978. DOI: https://doi.org/10.1016/j.jafr.2024.100978
CrossRef - Chaudhary, S., Chadha, M., Tyagi, B., Singh, K. Contribution of Sorghum and Finger Millets for Sustainable Food and Nutritional Security. In: Sustainable Food Systems (Volume I) SFS: Framework, Sustainable Diets, Traditional Food Culture & Food Production, Cham: Springer Nature Switzerland. 2024; 297-319. DOI: https://doi.org/10.1007/978-3-030-74768-8_10
CrossRef - Lazăr, N. N., Râpeanu, G., Iticescu, C. Mitigating eggplant processing waste’s environmental impact through functional food developing. Trends Food Sci Technol. 2024: 147:104414. DOI: https://doi.org/10.1016/j.tifs.2024.104414
CrossRef - Mala, T., Piayura, S., Itthivadhanapong, P. Characterization of dried pineapple (Ananas comosus) peel powder and its application as a novel functional food ingredient in cracker product. Future Foods. 2024; 9:100322. DOI: https://doi.org/10.1016/j.fufo.2024.100322
CrossRef - Bhatnagar, P., Gururani, P., Parveen, A., Gautam, P., Joshi, N. C., Tomar, M. S., Kumar, V. Algae: A promising and sustainable protein-rich food ingredient for bakery and dairy products. Food Chem. 2024; 441:138322. DOI: https://doi.org/10.1016/j.foodchem.2023.138322
CrossRef - Sánchez Gavilán, I., Velázquez Ybarzabal, D., de la Fuente, V., Cámara, R. M., Sánchez-Mata, M. C., Cámara, M. Valorization of Salicornia patula Duval-Jouve Young Shoots in Healthy and Sustainable Diets. Nutrients. 2024; 16(3):358. DOI: https://doi.org/10.3390/nu16030358
CrossRef - Flowers, T. J., Colmer, T. D. Plant salt tolerance: adaptations in halophytes. Ann Bot. 2015; 115(3):327-331. DOI: https://doi.org/10.1093/aob/mcu267
CrossRef - Rahman, M. M., Mostofa, M. G., Keya, S. S., Siddiqui, M. N., Ansary, M. M. U., Das, A. K., Tran, L. S. P. Adaptive mechanisms of halophytes and their potential in improving salinity tolerance in plants. Int J Mol Sci. 2021; 22(19):10733. DOI: https://doi.org/10.3390/ijms221910733
CrossRef - Lopes, M., Sanches-Silva, A., Castilho, M., Cavaleiro, C., Ramos, F. Halophytes as source of bioactive phenolic compounds and their potential applications. Crit Rev Food Sci Nutr. 2023; 63(8):1078-1101. DOI: https://doi.org/10.1080/10408398.2021.1959295
CrossRef - Salazar-López, N. J., González-Aguilar, G. A., Rouzaud-Sández, O., & Robles-Sánchez, M. Bioaccessibility of hydroxycinnamic acids and antioxidant capacity from sorghum bran thermally processed during simulated in vitro gastrointestinal digestion. J Food Sci Technol. 2018; 55:2021-2030. DOI: https://doi.org/10.1007/s13197-018-3116-z
CrossRef - Borgeat Meza, M., Luengo-Charath, X., Arancibia, M., Madrid, E. Pautas Éticas del Consejo de Organizaciones Internacionales de las Ciencias Médicas: avances y temáticas no resueltas en la actualización 2016. 2018; 18(2):e7208. DOI: https://doi.org/10.5867/medwave.2018.02.7208
CrossRef - Salazar-Lopez, N. J.; Loarca-Piña, G.; Campos-Vega, R.; Gaytán Martínez, M.; Morales Sánchez, E.; Esquerra-Brauer, J.M.; Robles Sánchez, M. The extrusion process as an alternative for improving the biological potential of sorghum bran: phenolic compounds and antiradical and anti-inflammatory capacity. Hindawi Evid Based Complementary Altern Med. 2016; 8387975. DOI: https://doi.org/10.1155/2016/8387975
CrossRef - Valenzuela-González, M.; Rouzaud-Sández, O.; Ledesma-Osuna, A. I.; Astiazarán-García, H.; Salazar-López, N. J.; Vidal-Quintanar, R. L.; Robles-Sánchez, M. Bioaccessibility of phenolic compounds, antioxidant activity, and consumer acceptability of heat-treated quinoa cookies. Food Sci Technol. 2022; 42, DOI: https://doi.org/10.1590/fst.43421
CrossRef - Salazar-López, N. J.; Astiacarán-García, H.; González-Aguilar, G. A.; Loarca-Piña, G.; Esquerra-Brauer, J. M.; Domínguez Avila, J. A.; Robles-Sánchez, M. Ferulic acid on glusose dysregulation, dyslipdemia, and inflammation in diet-induced obese rats: an integrated study. Nutrients. 2017; 9(7):675, DOI:https://doi.org/10.3390/nu9070675
CrossRef - Costa-Becheleni, F. R., Troyo-Diéguez, E., Ruiz-Hernández, A. A., Ayala-Niño, F., Bustamante-Salazar, L. A., Medel-Narváez, A., Robles-Sánchez, R. M. Determination of bioactive compounds and antioxidant capacity of the halophytes Suaeda edulis and Suaeda esteroa (Chenopodiaceae): An option as novel healthy agro-foods. AIMS Agric Food. 2024; 9(3):716-742, DOI: https://www.aimspress.com/article/doi/10.3934/agrfood.2024039
CrossRef - Lee, K. M.; Kalyani, D.; Tiwari, M. K.; Kim, T. S.; Dhiman, S. S.; Lee, J. K.; Kim, I. W. Enhanced enzymatic hydrolysis of rice straw by removal of phenolic compounds using a novel laccase from yeast Yarrowia lipolytica. Bioresour Technol. 2012; 123, 636-645. DOI: https://doi.org/10.1016/j.biortech.2012.07.066
CrossRef - Kolmogorov-Smirnov Test of Normality (Social Science Statistics). Available: https://www.socscistatistics.com/tests/kolmogorov/default.aspx
- Levene´s Homocedasticity Test of Similarity of Variances (Statistics Kingdom). Available: https://www.statskingdom.com/230var_levenes.html
- PAST (The Past of the Future) Versión 4.10 (mac). Available from: https://www.nhm.uio.no/english/research/infrastructure/past/
- Domínguez-Avila, J. A., Wall-Medrano, A., Velderrain-Rodríguez, G. R., Chen, C. Y. O., Salazar-López, N. J., Robles-Sánchez, M., González-Aguilar, G. A. Gastrointestinal interactions, absorption, splanchnic metabolism, and pharmacokinetics of orally ingested phenolic compounds. Food Funct. 2017; 8(1):15-38. DOI: 1039/C6FO01475E
CrossRef - Podsędek, A., Redzynia, M., Klewicka, E., Koziołkiewicz, M. Matrix effects on the stability and antioxidant activity of red cabbage anthocyanins under simulated gastrointestinal digestion. Biomed Res Int. 2014; 2014:365738. DOI: https://doi.org/10.1155/2014/365738
CrossRef - Shahidi, F., Yeo, J. Insoluble-bound phenolics in food. Molecules. 2016; 21(9):1216. DOI: https://doi.org/10.3390/molecules21091216
CrossRef - Zhang, B., Zhang, Y., Li, H., Deng, Z., Tsao, R. A review on insoluble-bound phenolics in plant-based food matrix and their contribution to human health with future perspectives. Trends Food Sci Technol. 2020; 105:347-362. DOI: https://doi.org/10.1016/j.tifs.2020.09.029
CrossRef - Borges, G., Lean, M. E., Roberts, S. A., Crozier, A. Bioavailability of dietary (poly) phenols: a study with ileostomists to discriminate between absorption in small and large intestine. Food Funct. 2013; 4(5):754-762. DOI: https://doi.org/10.1039/C3FO60024F
CrossRef - Gong, X., Hui, X., Wu, G., Morton, J. D., Brennan, M. A., Brennan, C. S. In vitro digestion characteristics of cereal protein concentrates as assessed using a pepsin-pancreatin digestion model. Food Res Int. 2022; 152:110715. DOI: https://doi.org/10.1016/j.foodres.2021.110715
CrossRef - McQuilken, S. The mouth, stomach, and intestines. Anaesth Intensive Care Med. 2024. DOI: https://doi.org/10.1016/j.mpaic.2024.01.006
CrossRef - Buitimea-Cantúa, N. E., Gutiérrez-Uribe, J. A., Serna-Saldívar, S. O. Phenolic–protein interactions: Effects on food properties and health benefits. J Med Food. 2018; 21(2):188-198. DOI: https://doi.org/10.1089/jmf.2017.0057
CrossRef - Amoako, D., Awika, J. M. Polyphenol interaction with food carbohydrates and consequences on availability of dietary glucose. Curr Opin Food Sci. 2016; 8:14-18. DOI: https://doi.org/10.1016/j.cofs.2016.01.010
CrossRef - Giuberti, G., Rocchetti, G., Lucini, L. Interactions between phenolic compounds, amylolytic enzymes and starch: An updated overview. Curr Opin Food Sci. 2020; 31:102-113. DOI: https://doi.org/10.1016/j.cofs.2020.04.003
CrossRef - Jakobek, L. Interactions of polyphenols with carbohydrates, lipids, and proteins. Food Chem. 2015; 175:556.567. DOI: https://doi.org/10.1016/j.foodchem.2014.12.013
CrossRef - Celep, E., İnan, Y., Akyüz, S., Yesilada, E. The bioaccessible phenolic profile and antioxidant potential of Hypericum perfoliatum after simulated human digestion. Ind Crops Prod. 2017; 109:717-723. DOI: https://doi.org/10.1016/j.indcrop.2017.09.032
CrossRef - Zhang, Q., Xing, B., Sun, M., Zhou, B., Ren, G., Qin, P. Changes in bio‐accessibility, polyphenol profile and antioxidants of quinoa and djulis sprouts during in vitro simulated gastrointestinal digestion. Food Sci Nutr. 2020; 8(8):4232-4241. DOI: https://doi.org/10.1002/fsn3.1718
CrossRef - Sousa, H. G. In vitro bioacessibility and bioactivity of phenolic compounds from halophyte plants. Universidad Nova de Lisboa. Nova School of Science & Technology. Department of Chemistry, 2022; Available: https://run.unl.pt/bitstream/10362/145155/1/Sousa_2022.pdf
- Muñoz-Bernal, Ó. A., Torres-Aguirre, G. A., Núñez-Gastélum, J. A., Rosa, L. A., Rodrigo-García, J., Ayala-Zavala, J. F., Álvarez-Parrilla, E. New approach to the interaction between Folin-Ciocalteu reactive and sugars during the quantification of total phenols. TIP Revista Especializada en Ciencias Químico-Biológicas. 2017; 20(2): 23-28. DOI: https://doi.org/10.1016/j.recqb.2017.04.003
CrossRef - Rodger, A., Sanders, K. Uv-visible absorption spectroscopy, bio-macromolecular applications. In: Encycl of spectroscopy and spectrometry Elsevier. 2017; 495-502. DOI: 1016/B978-0-12-803224-4.00108-4
CrossRef - Shraim, A. M., Ahmed, T. A., Rahman, M. M., Hijji, Y. M. Determination of total flavonoid content by aluminum chloride assay: A critical evaluation. Lwt. 2021; 150:111932. DOI: https://doi.org/10.1016/j.lwt.2021.111932
CrossRef - Goulas, V., Hadjisolomou, A. Dynamic changes in targeted phenolic compounds and antioxidant potency of carob fruit (Ceratonia siliqua) products during in vitro digestion. Lwt, 2019; 101:269-275. DOI: https://doi.org/10.1016/j.lwt.2018.11.003
CrossRef - Ma, Y., Yang, Y., Gao, J., Feng, J., Shang, Y., Wei, Z. Phenolics and antioxidant activity of bamboo leaves soup as affected by in vitroFood Chem Toxicol. 2020; 135: 110941. DOI: https://doi.org/10.1016/j.fct.2019.110941
CrossRef - Ketnawa, S., Reginio Jr, F. C., Thuengtung, S., Ogawa, Y. Changes in bioactive compounds and antioxidant activity of plant-based foods by gastrointestinal digestion: A review. Crit Rev Food Sci Nutr. 2022; 62(17): 4684-4705. DOI: https://doi.org/10.1080/10408398.2021.1878100
CrossRef - Parcheta, M., Świsłocka, R., Orzechowska, S., Akimowicz, M., Choińska, R., Lewandowski, W. Recent developments in effective antioxidants: The structure and antioxidant properties. Materials. 2021; 14(8):1984. DOI: https://doi.org/10.3390/ma14081984
CrossRef - Chen, J., Yang, J., Ma, L., Li, J., Shahzad, N., Kim, C. K. Structure-antioxidant activity relationship of methoxy, phenolic hydroxyl, and carboxylic acid groups of phenolic acids. Sci Rep. 2020; 10(1): 2611. DOI: https://doi.org/10.1038/s41598-020-59451-z
CrossRef - Chen, X., Li, H., Zhang, B., Deng, Z. The synergistic and antagonistic antioxidant interactions of dietary phytochemical combinations. Crit Rev Food Sci Nutr. 2022; 62(20): 5658-5677. DOI: https://doi.org/10.1080/10408398.2021.1888693
CrossRef - Shahidi, F., Zhong, Y. Measurement of antioxidant activity in food and biological systems. ACS Symposium Series eBooks. 2007; 956(4):36-66. DOI: 1021/bk-2007-0956.ch004
CrossRef - Nawar, W. W. Thermal degradation of lipids. J Agric Food Chem. 1969; 17(1):18-21. DOI: https://doi.org/10.1021/jf60161a012
CrossRef - Shahidi, F., Zhong, Y. Lipid oxidation and improving the oxidative stability. Chem Soc Rev. 2010; 39(11):4067-4079. DOI: https://doi.org/10.1039/B922183M
CrossRef - Shahidi, F., Zhong, Y. Measurement of antioxidant activity. J Funct Foods. 2015; 18:757-781. DOI: https://doi.org/10.1016/j.jff.2015.01.047
CrossRef - Acosta-Estrada, B. A., Gutiérrez-Uribe, J. A., Serna-Saldívar, S. O. Bound phenolics in foods, a review. Food Chem. 2014; 152:46-55. DOI: https://doi.org/10.1016/j.foodchem.2013.11.093
CrossRef - Waśkiewicz, A., Muzolf-Panek, M., Goliński, P. Phenolic content changes in plants under salt stress. In: Ecophysiology and Responses of Plants under Salt Stress, Ahmad, P., et al. Azooz, M., Prasad, M. Eds., Springer, New York, NY. 2013. DOI: https://doi.org/10.1007/978-1-4614-4747-4_11
CrossRef - Hajimehdipoor, H., Shahrestani, R., Shekarchi, M. Investigating the synergistic antioxidant effects of some flavonoid and phenolic compounds. Res J Phar. 2014; 1(3):35-40. Available from: https://www.rjpharmacognosy.ir/article_5776_609.html
- Rúa, J., de Arriaga, D., García-A., M. R., Busto, F., Del Valle, P. Binary combinations of natural phenolic compounds with gallic acid or with its alkyl esters: An approach to understand the antioxidant interactions. Eur Food Res Technol. 2017; 243:1211-1217. DOI: https://doi.org/10.1007/s00217-016-2838-2
CrossRef - Vuolo, M. M., Lima, V. S., Junior, M. R. M. Phenolic compounds: Structure, classification, and antioxidant power. Bioact Compd. 2019; 33-50. DOI: https://doi.org/10.1016/B978-0-12-814774-0.00002-5
CrossRef - Drawbridge, P. C., Apea-Bah, F., Hornung, P. S., Beta, T. Bioaccessibility of phenolic acids in Canadian hulless barley varieties. Food Chem. 2021; 358:129905. DOI: https://doi.org/10.1016/j.foodchem.2021.129905
CrossRef - Tomé-Sánchez, I., Martín-Diana, A. B., Peñas, E., Frias, J., Rico, D., Jiménez-Pulido, I., Martínez-Villaluenga, C. Bioprocessed wheat ingredients: Characterization, bioaccessibility of phenolic compounds, and bioactivity during in vitroFront Plant Sci. 2021; 12:790898. DOI: https://doi.org./10.3389/fpls.2021.790898
CrossRef - Quan, T. H., Benjakul, S., Sae-leaw, T., Balange, A. K., Maqsood, S. Protein–polyphenol conjugates: Antioxidant property, functionalities, and their applications. Trends Food Sci Technol. 2019; 91:507-517. DOI: https://doi.org/10.1016/j.tifs.2019.07.049
CrossRef - Cirillo, G., Curcio, M., Vittorio, O., Iemma, F., Restuccia, D., Spizzirri, U. G., Picci, N. Polyphenol conjugates and human health: a perspective review. Crit Rev Food Sci Nutr. 2016; 56(2):326-337. DOI: https://doi.org/10.1080/10408398.2012.752342
CrossRef - Bento-Silva, A., Patto, M. C. V., do Rosário Bronze, M. Relevance, structure, and analysis of ferulic acid in maize cell walls. Food Chem. 2018; 246:360-378. DOI: https://doi.org/10.1016/j.foodchem.2017.11.012
CrossRef - Miafo, A. P. T., Koubala, B. B., Kansci, G., Muralikrishna, G. Antioxidant properties of free and bound phenolic acids from bran, spent grain, and sorghum seeds. Cereal Chem. 2020; 97(6):1236-1243. DOI: https://doi.org/10.1002/cche.10348
CrossRef - Rodriguez-Mateos, A., Vauzour, D., Krueger, C. G., Shanmuganayagam, D., Reed, J., Calani, L., Crozier, A. Bioavailability, bioactivity, and impact on health of dietary flavonoids and related compounds: an update. Arch Toxicol. 2014; 88:1803-1853. DOI: https://doi.org/10.1007/s00204-014-1330-7
CrossRef - Velderrain-Rodríguez, G. R., Palafox-Carlos, H., Wall-Medrano, A., Ayala-Zavala, J. F., Chen, C. O., Robles-Sánchez, M., González-Aguilar, G. A. Phenolic compounds: their journey after intake. Food Func. 2014; 5(2):189-197. DOI: https://doi.org/10.1039/C3FO60361J
CrossRef - Gao, J., Yu, H., Guo, W., Kong, Y., Gu, L., Li, Q., Wang, Y. The anticancer effects of ferulic acid is associated with induction of cell cycle arrest and autophagy in cervical cancer cells. Cancer Cell Int. 2018; 18:1-9. DOI: https://doi.org/10.1186/s12935-018-0595-y
CrossRef - Adefegha, S. A., Oboh, G., Ejakpovi, I. I., Oyeleye, S. I. Antioxidant and antidiabetic effects of gallic and protocatechuic acids: a structure–function perspective. Comp Clin Pathol. 2015; 24:1579-1585. DOI: https://doi.org/10.1007/s00580-015-2119-7
CrossRef - Petropoulos, S. A., Karkanis, A., Martins, N., Ferreira, I. C. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci Technol. 2018; 74:69-84. DOI: https://doi.org/10.1016/j.tifs.2018.02.006
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






