
Introduction
Hypercholesterolemia or excess blood cholesterol, is one of the risk factors for atherosclerosis, which promotes cardiovascular diseases. Cardiovascular diseases (CVD) lead to high mortality worldwide, with about 18.6 million deaths in 2019.1 Sources of cholesterol in the human body are from diet and through de-novo synthesis in the liver. High levels of blood cholesterol, especially low-density lipoprotein cholesterol (LDL), can cause hypercholesterolemia. This condition is associated with an increased risk of atherosclerosis.2 There are several strategies to treat hypercholesterolemia, including inhibiting 3-hydroxy-3-methylglutaryl coenzyme A reductase (HMGR) activity and using bile acid-binding agents. HMGR is one of the enzymes essential to the synthesis of cholesterol in the liver. Inhibiting HMGR reduces cholesterol synthesis and increases the number of LDL receptors, which helps lower LDL cholesterol in the blood. Bile acid-binding substances reduce bile acid levels in the enterohepatic circulation, leading to lower blood cholesterol as cholesterol is converted to bile acids.3 Hypercholesterolemia and atherosclerotic risk factors increase the generation of reactive oxygen species (ROS) in the vessel walls, causing oxidative stress.4
It is well known that many naturally derived active compounds, including peptides, have great impact on the prevention or treatment of various disorders such as CVD, cancer, and diabetes. Bioactive peptides are potential therapeutic components in the pharmaceutical and biotechnology industries. They exhibit a strong affinity for target molecules, are stable against proteolytic degradation, safe, and multifunctional5,6 Bioactive peptides can be obtained by the enzymatic hydrolysis process of dietary proteins from plant, marine, and animal sources.7 Enzymatic hydrolysis is considered more advantageous than the chemical hydrolysis process because it uses mild conditions and is more accurate in the cleavage of peptide bonds.8 The types of enzymes employed in the hydrolysis process determine the diversity of physicochemical and biological properties of hydrolysate proteins and peptides.9 Each protease enzyme targets specific peptide bonds, producing peptides with varying molecular weights, sizes, and amino acid sequences, resulting in diverse properties.6 Many studies have demonstrated the various biological activities of hydrolysate protein and peptides, including antioxidant and anti-hypercholesterolemic. Cowpea,10 hempseed,9,11 and cumin seed 3,6 are among the plant-derived protein with in-vitro antioxidant and hypocholesterolemic capacity.
Melon (Cucumis melo L.) is a popular fruit that belongs to the Cucurbitaceae family and it is one of the most consumed fruits worldwide with high economic value. However, by-products of the fruit such as seeds, are usually discarded during industrial processing or consumption. In several countries, melon seeds are also consumed after being roasted or are utilized as flavoring agents for traditional dishes and desserts. 12–14 Melon seeds possess a high content of protein (14.91%- 36.3%)15, however, there is a lack of exploration regarding the biological activities of melon seed protein. Determining the biological effects of melon seeds could increase their value and usage, particularly in the pharmaceutical and food industries.
In vitro studies by Siddeeg et al.16 reported that the protein of Cucumis melo var. tibish seed, when sequentially digested with trypsin and pepsin, exhibits antioxidant activity. However, research on the bioactivities of melon (Cucumis melo L.) seed protein (MSP) hydrolysate is currently limited. Specifically, no previous reports are available on the hypocholesterolemic activity of MSP hydrolysate. This study aims to investigate the antioxidant and hypocholesterolemic activities of MSP hydrolysate using three different proteases, i.e., pepsin, thermolysin, and trypsin, which are commonly used to produce bioactive peptides. Our findings highlight the potential of MSP hydrolysates as nutraceutical agents, identifying their antioxidant and hypocholesterolemic properties for versatile applications in the food and pharmaceutical sectors.
Materials and Methods
Material
Melon (Cucumis melo L.) fruit was obtained from the local market, and the seeds were removed manually from the fruit. The identification of the plant material was carried out in the Herbarium Depokensis (UIDEP) at the Department of Biology, Indonesia University. Protease enzymes used were pepsin (1:10,000 or 10,000 NFU/mg) from Himedia, trypsin (0.2 Anson units/g) from Himedia, and thermolysin (30-350 units/mg) from Sigma Aldrich.
Preparation of Melon Protein Hydrolysate
Melon seeds were dried at 40 oC and then milled to obtain seed flour. Preparation to obtain protein from melon seed was conducted by conventional alkaline extraction-isoelectric precipitation method as described by Marques et al.10 The flour of the seeds was defatted using soxhlet extraction with n-hexane for 5 h at 60 – 70 °C and the defatted flour was suspended in deionized water with a ratio of 1:10 (w/v). After adding 1 M NaOH to adjust the pH of the mixture to 9.3, it was agitated using a magnetic stirrer at 500 rpm for 2 h at room temperature. Then the slurry was centrifuged at 5500 ×g at a temperature of 4 oC for 15 min, and the extraction process was carried out once more after the residue was suspended in water. All of the supernatants were mixed and precipitated at pH 4.4 with the addition of 1 M HCl. The precipitate was separated by centrifugation at 5500 ×g at 4 oC for 15 min. The precipitate obtained was melon seed protein isolate, then washed with water and the pH adjusted to 7 by adding 1 M NaOH. The melon seed protein isolate was dried using a freeze-dryer (Buchi Lyovapor L-300). The next step was the hydrolysis process of the freeze-dried MSP, which was conducted with the procedure described by He et al.5 with minor modifications. MSP was dissolved (5% w/v protein basis) in deionized water in a reaction vessel with a stirrer. Then, the samples were digested for 4 h by three different types of enzymes at an enzyme-to-substrate (E/S) ratio of 1:50 (w/w). Temperature and pH were adjusted to the optimum conditions for each enzyme. Trypsin, at 37 oC and pH 8; pepsin, 37 oC and pH 2; and thermolysin, 55 oC and pH 8. During the hydrolysis, the pH of solution was kept constant. Reactions were stopped to inactivate the enzyme by heating the solution in a boiling water bath for about 20 min and then centrifuged at 5500 xg at 4 oC for 30 min. The supernatant was collected as hydrolysate and stored at -20 oC for further analysis. The protein content of the hydrolysate was determined using the Bradford method.17
Amino acid composition Determination
Analysis of amino acids was performed by acid hydrolysis and using the UPLC system with guidelines according to System Guide of Waters Acquity UPLC H Class and H Class Bio amino Acid Analysis.18 Each sample of 1 μL was injected into column AccQ.Tag Ultra C18 1.7 μm (2.1 x 100 mm), at 49 °C with PDA detection at 260 nm wavelength. The mobile phase had a flow rate of 0.5 mL/min and the following composition: mobile phase A was AccQ.Tag Ultra amino acid analysis eluent A; mobile phase B was AccQ.Tag Ultra amino acid analysis 10% (in water), mobile phase C was Aquabidest grade HPLC and mobile phase D was AccQ.Tag Ultra amino acid analysis eluent B. While tryptophan (Trp) content was analyzed after alkaline hydrolysis using High Performance Liquid Chromatography (HPLC) in compliance with AOAC official method 988.15. Each sample of 15 μL was injected into column RP-18.5 μm, 250 mm x 4.0 mm, at ambient temperatures. The mobile phase flow rate was 1.5 mL/min, with phase A being 0.0085 M sodium acetate (pH 4) and phase B methanol. The detector was a PDA at 280 nm wavelength.
Antioxidant Activity Measurement
ABTS radical scavenging activity assay
The ABTS (2,2’-azinobis-(3ethylbenzothiazoline)-6-sulfonic acid) radical scavenging properties of hydrolysates were conducted using the procedure from Zhuang et al.19 with slight modifications. The radical cation of ABTS was produced by mixing 5 mL of 7 mM ABTS stock solution with 85 µL of 2.45 mM potassium persulfate. The mixture was incubated at room temperature in the dark for 14–16 h. Then 0.2 M of PBS pH 7.4 was used to dilute the ABTS radical solution until its absorbance was 0.7 at 734 nm. 2 mg/mL of hydrolysate solution was mixed with the diluted ABTS radical solution (1:4). The absorbance of this mixture was recorded at 734 nm using a BioTek Instrument Synergy H1MF microplate spectrophotometer after incubating for 6 min. The percentage of radical inhibition by the sample was calculated using the formula below, with PBS as a blank:
% ABTS• scavenging activity = [(blank absorbance – sample absorbance)/( blank absorbance)] x 100%
DPPH radical scavenging activity assay
To measure DPPH (2,2-diphenyl-1-picrylhydrazyl) radical scavenging properties, the method by Ben Khaled et al.20 was followed with minor modifications. 500 µL of hydrolysate (9 mg/mL) in 0.1 M phosphate buffer pH 7 was added to 375 µL of 90% ethanol and 125 µL of 0.4 mM DPPH in ethanol. After an hour of incubation at 37 °C, the absorbance was recorded at 517 nm using a BioTek Instrument Synergy H1MF microplate spectrophotometer. The percent inhibition of DPPH radicals by the hydrolysate was calculated using the formula below:
% DPPH• scavenging activity = [(blank absorbance – sample absorbance)/( blank absorbance)] x 100%
Ferrous ion chelating ability assay
The activity of hydrolysate to chelate ferrous ions was determined by following the method of Yuan et al.21 with slight modifications. 1 mL of hydrolysate (9 mg/mL) in sodium acetate buffer pH 5 was mixed with 100 µL of 2 mM FeCl2.4H2O, then incubated at 37 oC for 30 min. After adding 200 µL of 5 mM ferrozine to the mixture, it was incubated for 10 min at room temperature. The absorbance of the sample was read using BioTek Instrument Synergy H1MF microplate spectrophotometer at 562 nm. The chelating activity was calculated using the formula as follows:
% Chelating activity = [(blank absorbance – sample absorbance)/(blank absorbance)] x 100%
Hypocholesterolemic Activity
HMG-CoA Reductase Inhibition Assay
The HMG-CoA reductase (HMGR) inhibition test was conducted using the Sigma-Aldrich® HMG-CoA reductase assay kit CS 1090-1KT. In summary, each hydrolysate sample with NADPH, substrate (HMG-CoA), and buffer was loaded into a 96-well plate. The reactions were initiated by HMG-CoA reductase addition in each well, and the absorbance was read at 340 nm. The specific activity of the HMGR enzyme was interpreted as μmol of oxidized NADPH/min/mgP. The inhibitory capacity of the hydrolysates was determined by reading the decrease in absorbance at 340 nm for 10 min, and this absorbance was directly related to the activity of the enzymes. The results were given as a percentage of the HMGR enzyme’s control specific activity without hydrolysate or pravastatin. Pravastatin was a positive control, and the HMGR inhibitory activity of the hydrolysate was calculated as follows:
% Inhibition = [(blank HMGR enzyme activity – HMGR enzyme activity in the precense hydrolysate)/(blank HMGR enzyme activity)] x 100%
Bile Acid Binding Activity
The determination of the bile-acid binding ability was performed using the method of Yoshie-Stark 22. In brief 900 µl of bile acid solution i.e., sodium deoxycholate 2 mM was added to 100 µL of hydrolysate sample (10 mg/mL) in phosphate buffer (0.1 mM pH 7). After incubation at 37 oC for 2 h, each sample was centrifuged at 12000 rpm 4 oC for 10 min. The result supernatant was diluted, and the bile acid content in sample was analyzed following Bile Acid kit MAK309 Sigma Aldrich instructions. All samples were run in duplicate and analyzed fluorescence spectrophotometrically at λex 530 nm and λem 585. The binding activity was determined using the formula as follows:
% Bile acid binding activity = [(bile acid in blank – bile acid in sample)/( bile acid in blank)] x 100%
Statistical analysis
All tests score are reported as mean ± standard deviation (SD). The one way analysis of variance (ANOVA) test was used in IBM SPSS Statistics version 23 for statistical analysis. A significant difference in means was identified between the samples at p < 0.05.
Results and Discussion
Protein Content and Amino Acid Composition
The soluble protein contents of MSP hydrolysate are 2.28 mg/mL, 2.36 mg/mL, and 4.38 mg/mL for pepsin, thermolysin and trypsin-treated hydrolysate respectively (Figure 1). According to Kim et al.23 differences in protein content may arise from the enzymes selected for hydrolysis. Despite the many factors that could affect hydrolysate protein content, the enzyme used for hydrolysis plays a crucial role in determining peptide bond cleavage patterns, which correlate with protein levels in the hydrolysate. Their study23 demonstrated varying protein content in perilla seed meal protein hydrolysates depending on the enzyme used. In addition, Alashi et al.24 have also reported that canola meal protein hydrolysates from trypsin and chymotrypsin digestion had the highest levels of protein content compared to alcalase, pancreatin, and pepsin hydrolysates.
The amino acid compositions of MSP hydrolysates are summarized in Table 1. The amino acids with the highest levels in all samples of hydrolysate are glutamic acid (0.505-0.977 g/100 g sample) and arginine (0.682-0.801 g/100 g sample), which is in accordance with the previous study reported by Siddeeg et al.25 Followed by aspartic acid (0.244-0.49 g/100 g sample), phenylalanine (0.307-0.359 g/100 g sample), and leucine (0.285-0.387 g/100 g sample). Dash and Gosh26 reported that major amino acids found in Cucurbitaceae seed protein hydrolysate are glutamic acid, aspartic acid, and arginine.
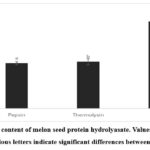 |
Figure 1: Protein content of melon seed protein hydrolyasate. Values are expressed as means ± SD. Various letters indicate significant differences between means (p < 0.05) |
Protein hydrolysates, obtained through enzymatic hydrolysis, contain amino acids and bioactive peptides, valued for their nutraceutical benefits and critical roles in vital human physiological processes.27 The composition of amino acid and peptide sequences plays a crucial role in determining the bioactivity of the hydrolysate.28 Aromatic amino acids such as tyrosine and phenylalanine can donate electrons to convert free radicals into stable molecules, thereby enhancing biological activity29. The negatively charged acidic amino acids have excess electrons that can be donated to quenching free radicals. Therefore, they can act as antioxidants.30,31 Hydrophobic amino acids have the ability to donate protons and electrons, and enhance the lipid solubility of peptides, potentially increasing their antioxidant activity.27,29 Furthermore, Maestri et al.32 suggested that hydrophobic bioactive peptides containing leucine, tryptophan, and tyrosine are reported to exhibit better hypocholesterolemic activity. Prados et al.33 revealed that thirty-three peptides identified in the olive seed hydrolysate were found to be rich in hydrophobic amino acids, potentially contributing to their hypolipidemic properties. Several studies have confirmed that peptides from food protein hydrolysates with antioxidant properties exhibit additional biological activities, including hypocholesterolemic effects.34
In the present study, the hydrolysates from thermolysin digestion have high hydrophobic amino acids (Table 1) such as, leucine, valin , methionine, prolin, and phenylalanine. The hydrolysates also contained more aromatic amino acids, as well as the negatively charged acidic amino acids (Table 1). These results suggest that the hydrolysate produced by thermolysin may have stronger antioxidant and hypocholesterolemic properties compared to those produced by pepsin and trypsin.
Table 1: Amino acid composition of melon seed protein hydrolysates (g/100g sample)
| Amino Acids | Pepsin | Thermolysin | Trypsin |
| Serine | 0.214 | 0.306 | 0.228 |
| Glutamic acid | 0.505 | 0.977 | 0.562 |
| Phenylalanine | 0.307 | 0.359 | 0.350 |
| Isoleucine | 0.161 | 0.185 | 0.153 |
| Valine | 0.191 | 0.219 | 0.176 |
| Alanine | 0.148 | 0.252 | 0.143 |
| Arginine | 0.682 | 0.854 | 0.801 |
| Glycine | 0.200 | 0.274 | 0.211 |
| Lysine | 0.086 | 0.162 | 0.079 |
| Aspartic acid | 0.244 | 0.490 | 0.245 |
| Leucine | 0.285 | 0.397 | 0.269 |
| Tyrosine | 0.157 | 0.185 | 0.179 |
| Proline | 0.141 | 0.201 | 0.136 |
| Threonine | 0.183 | 0.225 | 0.192 |
| Histidine | 0.132 | 0.138 | 0.148 |
| Tryptophan | 0.044 | 0.902 | 0.049 |
| Cystine | 0.056 | 0.318 | 0.042 |
| Methionine | 0.009 | 0.119 | 0.012 |
| EAA* | 1.401 | 1.790 | 1.431 |
| HAA | 1.459 | 1.844 | 1.463 |
| NCAA | 0.749 | 1.468 | 0.807 |
| PCAA | 0.901 | 1.156 | 1.029 |
| AAA | 0.509 | 0.635 | 0.579 |
*Essential amino acids (EAA)- histidine, lysine, methionine, leucine, threonine, tryptophan, phenylalanine, isoleucine, valine; Hyrdrophobic amino acid (HAA) – alanine, proline, cysteine, valine, methionine, isoleucine, leucine, tyrosine, and phenylalanine; Negatively charged amino acid (NCAA) – aspartic acid, glutamic acid; Positively charged amino acid (PCAA)- histidine, lysine, arginine; Aromatic amino acid (AAA) – tryptophan, tyrosine, phenylalanine
Antioxidant Activity
Radical Scavenging Acitivty
ABTS radical scavenging activity of hydrolysate at concentration of 2 mg/mL is shown in Figure 2A. The ABTS radical scavenging method can be used to determine the ability of both lipophilic and hydrophilic molecules to function as antioxidants. In this study, hydrolysate digested with thermolysin exhibited greatest ability against ABTS. The scavenging activity of thermolysin-treated hydrolysate was 47.38 ± 1.51% and significantly different (p < 0.05) from trypsin (37.65 ± 0.9%) and pepsin (35.27 ± 1.62%). This result indicates that the type of enzyme used for hydrolysis affected the activity to scavenge ABTS radical. A study by Garcia et al.35 also found that hydrolysates of cherry seeds digested with thermolysin exhibit the highest ABTS radical scavenging activity compared to alcalase and flavorzyme. Zheng et al.36 suggested residues such as tyrosine, tryptophan, or cysteine are required to scavenge ABTS. The mechanism by which antioxidants neutralize ABTS radicals is through hydrogen and electron donors.28 Amino acids with SH, NH, and OH side chains, such as tyrosine, cysteine, and tryptophan, are considered to play an important role in scavenging ABTS radicals due to the presence of a labile hydrogen atom.37
DPPH is a relatively stable free radical that can accept an electron or hydrogen. The DPPH radical scavenging activity of MSP hydrolysate at a concentration of 9 mg/mL is shown in Figure 2B. MSP digested by thermolysin has the strongest capacity to scavenge DPPH radical (21.44 ± 0.37%) followed by pepsin (20.81 ± 0.99%) and trypsin (19.02 ± 0.63%). It was observed no significant difference (p > 0.05) between the thermolysin and pepsin-treated hydrolysate against DPPH radical but compared to trypsin-treated hydrolysate was significantly different (p <0.05). Our result is in line with previous work by Alashi et al.24 who reported the canola meal protein hydrolysate of pepsin was more active against DPPH radicals than the hydrolysate treated with trypsin. Moreover, Sun, et al.38 revealed the red algae protein hydrolysate of pepsin has a greater ability to inhibit DPPH radicals compared to trypsin. As reported by Liao et al.37 phenylalanine, proline, histidine and cysteine may serve as DPPH radical scavengers.
 |
Figure 2: Free radical scavenging activities of different enzymatic hydrolysates of MSP against ABTS radicals (A) and DPPH radicals (B). All data were presented as the mean ± SD. Significant differences (p < 0.05) are indicated by values with distinct letters. |
Metal (Ferrous) Ion Chelating Activity
Ferrous ions (Fe2+) can act as prooxidants due to being involved in the Fenton and Haber-Weiss reaction which produce hydroxyl radicals that can damage biomolecules such as DNA and protein cellular. In addition, transition metals such as iron and copper stimulate the lipid peroxidation process.39 Therefore metal chelation activity also relates to the antioxidant properties of bioactive compounds. The ferrous ion chelating activity of MSP hydrolysates is displayed in Figure 3.
The results observed that hydrolysates obtained by three proteases have strong chelating activity toward Fe2+ at the concentration of 9 mg/mL. The chelating activity exhibited significant differences (p < 0.05) among each three hydrolysates, with trypsin having the most potent activity (70.04 ± 2.40%), followed by thermolysin (56.22 ± 0.18%) and pepsin (25.59 ± 1.40%). In agreement with the current result, several works have also reported that trypsin-treated hydrolysate had the highest activity in binding ferrous ions.40–42 Trypsin demonstrated a much higher capacity to produce ferrous-chelating peptides of MSP might be due to the specificity of this enzyme to cleavage arginine or lysine residue at the C-terminal of the peptide bond.43 According to Tian et al. 44, positively charged amino acids such as lysine, arginine, and histidine have metal-binding ability due to the presence of the nitrogen atom. Moreover, Zhang et al.45 revealed that mung bean peptides chelate ferrous ions mainly through amino, carboxyl, and imidazole groups of amino acids.
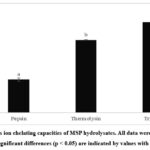 |
Figure 3: Ferrous ion chelating capacities of MSP hydrolysates. All data were presented as the mean ± SD. Significant differences (p < 0.05) are indicated by values with distinct letters. |
In general, the hydrolysates from three types of enzymes displayed significant differences (p <0.05) in activity to inhibit free radicals and to chelate metal ions. The findings obtained suggest that antioxidant capacity of MSP hydrolysates depends on the enzyme specificity and activity. Proteases with different specificities will produce peptides with different amino acid compositions, sequence, and molecular weights. Samaranayaka and Li-Chan46 suggested that antioxidant potency of peptides depend on amino acid composition, sequence, and molecular size.
Hypocholesterolemic Activity
HMG CoA Reductase Inhibition
The in-vitro investigation of the hypocholesterolemic activity of enzymatic hydrolysates from melon seed protein in this current study involved the HMGR inhibitory activity and bile acid binding capacity assay. Figure 4. shows the activities of HMGR inhibition by the three enzymatic hydrolysates with pravastatin used as a positive control. All the hydrolysates possessed inhibitory activity against HMGR at concentration of 2 mg/mL and are listed in decreasing order: thermolysin (68.25 ± 3.60%), pepsin (57.95 ± 5,11%) and trypsin (44.55 ± 5.99%). The hyrolysate of thermolysin demonstrated most potent inhibitor for HMGR and ANOVA test confirms that there were no significant differences (p > 0.05) between the inhibitory activity of thermolysin-treated hydrolysate and control positive pravastatin (81.67 ± 0.65%). While trypsin hydrolysate showed significantly (p < 0.05) lower activity compared to thermolysin and pepsin.
In accordance with our current study, several in vitro investigations have also confirmed the inhibitory activity of peptide fractions and hydrolysate from food and plant protein against HMGR. Prados et al.33 demonstrated that olive seed protein hydrolysate from alcalase could inhibit HMGR by 16.8 and 40% at the concentrations of 3.1 mg/mL and 40 mg/mL, respectively. Lupin peptide mixture of pepsin and trypsin at a concentration of 2.5 mg/mL1 could reduce HMGR activity by 17 and 61%, respectively, as reported by Lammi, et al.47 One of the inhibitory mechanisms of HMGR activity by peptides is through competitive inhibition of amino acid residues in the catalytic site of HMGR. Interactions between amino acids of the peptide from lupin with the active site of HMGR through electrostatic, hydrogen bond, salt bridge and hydrophobic interactions.48,49 In this present study, thermolysin and pepsin have a better ability to interfere with HMGR compared to trypsin, which might be due to peptides from trypsin digestion having positively charged amino acids such as arginine or lysine at the C-terminal. These amino acids are less unfit for the active site of HMGR, which is also surrounded by some positively charged residues.49
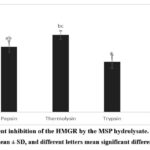 |
Figure 4: Percent inhibition of the HMGR by the MSP hydrolysate. The data were shown as the mean ± SD, and different letters mean significant differences (p < 0.05). |
Bile Salt Binding Capacity
For Figure 5 show sodium deoxycholate binding ability of the hydrolysates at concentration of 10 mg/mL. Among the three hydrolysates, thermolysin exhibited the highest bile acid-binding capacity (19.91 ± 1.43%), followed by pepsin (13.65 ± 0.98%) and trypsin (8.34 ± 0.11%). Our observed result are consistent with Mohan and Udenigwe50 who reported that hydrolysate of casein at concentration of 10 mg/mL have 12.8% bile acid binding capacity. Yoshie-Stark and Wäsche22 reported that soybean protein and its hydrolysate (10 mg/mL) of pepsin and pepsin/pancreatin have sodium deoxycholate binding capacity in a range of 12.6% – 15.0%, while lupin and its hydrolysate was 12.8% – 69.5%. Bile acid is an amphipathic molecule that consists of both a hydrophobic part due to the steroid core with hydrogen and methyl groups and a hydrophilic part with hydroxyl and carboxyl groups. These groups are considered to contribute to binding other molecules.3 Previous in silico study by Ngoh et al.51 described that most amino acids of peptides derived from pinto bean that interact with bile acid are hydrophobic amino acids such as proline, leucine, glycine, phenylalanine, alanine, and methionine. Moreover, Siow, et al3 reported that amino acids of peptides from cumin seed can bind bile acid molecules through hydrophobic and hydrophilic interactions.
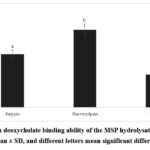 |
Figure 5: Sodium deoxycholate binding ability of the MSP hydrolysate. The data were shown as the mean ± SD, and different letters mean significant differences (p < 0.05). |
Conclusion
In conclusion, this present work displayed that MSP hydrolysates have in-vitro antioxidant and hypocholesterolemic properties. The biological capacity of hydrolysates varied depending on the type of protease used for protein digestion. The melon seed hydrolysate obtained using thermolysin enzyme preparation showed better abilities to inhibit HMGR, bind bile acid and scavenge free radicals compared to pepsin and trypsin. The hydrolysate from trypsin digestion has the highest activity in chelating metal ions. These findings suggest that the antioxidant and hypocholesterolemic properties of MSP hydrolysate might be useful in the preparation of nutraceuticals and in the development of pharmaceutical peptide-based products. In addition, melon seed may potentially be used as a raw material to produce bioactive ingredients with health benefits that could contribute to making this underutilized part of fruit more valuable. In future research, it would be beneficial to explore a variety of enzymes to build on the current study. Additionally, conducting in vivo studies will be necessary for improving these findings.
Acknowledgement
The author would like to thank the scholarship providers, namely the Indonesia Funds for Education (LPDP) and Center for Higher Education Funding (BPPT) for their support in carrying out the doctoral program in chemistry.
Funding Source
This research was funded by the scholarship providers, namely the Indonesia Funds for Education (LPDP) and the Center for Higher Education Funding (BPPT), with decree number: 0625/J5.2.3./BPI.06/10/2021.
Conflict of Interest
The authors declare no conflict of interest.
Authors’ Contribution
Deasy Natalia Botutihe: conceptualization, study design, collection of data, analysis of results, manuscript preparation, writing and editing.
Sumi Hudiyono: study design, supervision, and manuscript preparation.
Endang Saepudin: study design, supevision, and manuscript preparation.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal, subjects, or any material that requires ethical approval
References
- Roth GA, Mensah GA, Johnson CO, et al. Global Burden of Cardiovascular Diseases and Risk Factors, 1990-2019: Update From the GBD 2019 Study. J Am Coll Cardiol. 2020;76(25):2982-3021. doi:10.1016/j.jacc.2020.11.010
CrossRef - Collado A, Domingo E, Piqueras L, Sanz MJ. Primary hypercholesterolemia and development of cardiovascular disorders: Cellular and molecular mechanisms involved in low-grade systemic inflammation and endothelial dysfunction. International Journal of Biochemistry and Cell Biology. 2021;139. doi:10.1016/j.biocel.2021.106066
CrossRef - Siow HL, Choi SB, Gan CY. Structure–activity studies of protease activating, lipase inhibiting, bile acid binding and cholesterol-lowering effects of pre-screened cumin seed bioactive peptides. J Funct Foods. 2016;27:600-611. doi:10.1016/j.jff.2016.10.013
CrossRef - Kattoor AJ, Pothineni NVK, Palagiri D, Mehta JL. Oxidative Stress in Atherosclerosis. Curr Atheroscler Rep. 2017;19(11). doi:10.1007/s11883-017-0678-6
CrossRef - He R, Malomo SA, Alashi A, Girgih AT, Ju X, Aluko RE. Purification and hypotensive activity of rapeseed protein-derived renin and angiotensin converting enzyme inhibitory peptides. J Funct Foods. 2013;5(2):781-789. doi:10.1016/j.jff.2013.01.024
CrossRef - Siow HL, Gan CY. Extraction, identification, and structure-activity relationship of antioxidative and α-amylase inhibitory peptides from cumin seeds (Cuminum cyminum). J Funct Foods. 2016;22:1-12. doi:10.1016/j.jff.2016.01.011
CrossRef - Siow HL, Gan CY. Extraction of antioxidative and antihypertensive bioactive peptides from Parkia speciosa seeds. Food Chem. 2013;141(4):3435-3442. doi:10.1016/j.foodchem.2013.06.030
CrossRef - Villamil O, Váquiro H, Solanilla JF. Fish viscera protein hydrolysates: Production, potential applications and functional and bioactive properties. Food Chem. 2017;224:160-171. doi:10.1016/j.foodchem.2016.12.057
CrossRef - Girgih AT, Udenigwe CC, Aluko RE. In Vitro Antioxidant Properties of Hemp Seed ( Cannabis sativa L.) Protein Hydrolysate Fractions. J Am Oil Chem Soc. 2011;88(3):381-389. doi:10.1007/s11746-010-1686-7
CrossRef - Marques MR, Soares Freitas RAM, Corrêa Carlos AC, Siguemoto ÉS, Fontanari GG, Arêas JAG. Peptides from cowpea present antioxidant activity, inhibit cholesterol synthesis and its solubilisation into micelles. Food Chem. 2015;168(March):288-293. doi:10.1016/j.foodchem.2014.07.049
CrossRef - Zanoni C, Aiello G, Arnoldi A, Lammi C. Hempseed Peptides Exert Hypocholesterolemic Effects with a Statin-Like Mechanism. J Agric Food Chem. 2017;65(40):8829-8838. doi:10.1021/acs.jafc.7b02742
CrossRef - Rolim PM, Fidelis GP, Padilha CEA, Santos ES, Rocha HAO, Macedo GR. Phenolic profile and antioxidant activity from peels and seeds of melon (Cucumis melo L. var. reticulatus) and their antiproliferative effect in cancer cells. Brazilian Journal of Medical and Biological Research. 2018;51(4). doi:10.1590/1414-431×20176069
CrossRef - Bouaziz A, Djidel S, Bentaher A, Khennouf S. Polyphenolic content, Antioxidant and Anti-inflammatory activities of Melon (Cucumis melo L. var. inodorus) Seeds. Journal of Drug Delivery and Therapeutics. 2020;10(2-s):22-26. doi:10.22270/jddt.v10i2-s.4022
CrossRef - Rabadán A, Antónia Nunes M, Bessada SMF, Pardo JE, Beatriz Oliveira MPP, Álvarez-Ortí M. From by-product to the food chain: Melon (cucumis melo l.) seeds as potential source for oils. Foods. 2020;9(10). doi:10.3390/foods9101341
CrossRef - Silva MA, Albuquerque TG, Alves RC, Oliveira MBPP, Costa HS. Melon (Cucumis melo L.) by-products: Potential food ingredients for novel functional foods? Trends Food Sci Technol. 2020;98(April):181-189. doi:10.1016/j.tifs.2018.07.005
CrossRef - Siddeeg A, Xu Y, Jiang Q, Al-Farga A, Wenshui X. Influence of Enzymatic Hydrolysis on the Nutritional, Functional and Antioxidant Properties of Protein Hydrolysates Prepared from Seinat (Cucumis melo var. tibish) Seeds “Influence of Enzymatic Hydrolysis on the Nutritional, Functional and Antioxidant Pro. Journal of Food and Nutrition Research. 2015;3(4):259-266. doi:10.12691/jfnr-3-4-5
- Bradford MM. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Vol 72.; 1976.
CrossRef - Technical Communications Department W. ACQUITY UPLC H-Class and H-Class Bio Amino Acid Analysis System Guide Revision B.; 2012. www.waters.com.
- Zhuang H, Tang N, Yuan Y. Purification and identification of antioxidant peptides from corn gluten meal. J Funct Foods. 2013;5(4):1810-1821. doi:10.1016/j.jff.2013.08.013
CrossRef - Ben Khaled H, Ktari N, Ghorbel-Bellaaj O, Jridi M, Lassoued I, Nasri M. Composition, functional properties and in vitro antioxidant activity of protein hydrolysates prepared from sardinelle (Sardinella aurita) muscle. J Food Sci Technol. 2014;51(4):622-633. doi:10.1007/s13197-011-0544-4
CrossRef - Yuan B, Zhao C, Cheng C, et al. A peptide-Fe(II) complex from Grifola frondosa protein hydrolysates and its immunomodulatory activity. Food Biosci. 2019;32. doi:10.1016/j.fbio.2019.100459
CrossRef - Yoshie-Stark Y, Wäsche A. In vitro binding of bile acids by lupin protein isolates and their hydrolysates. Food Chem. 2004;88(2):179-184. doi:10.1016/j.foodchem.2004.01.033
CrossRef - Kim JM, Yoon KY. Functional properties and biological activities of perilla seed meal protein hydrolysates obtained by using different proteolytic enzymes. Food Sci Biotechnol. 2020;29(11):1553-1562. doi:10.1007/s10068-020-00810-x
CrossRef - Alashi AM, Blanchard CL, Mailer RJ, et al. Antioxidant properties of Australian canola meal protein hydrolysates. Food Chem. 2014;146:500-506. doi:10.1016/j.foodchem.2013.09.081
CrossRef - Siddeeg A, Xu Y, Jiang Q, Xia W. Physicochemical and functional properties of flour and protein isolates extracted from seinat (Cucumis melo var. tibish) seeds. Food Sci Biotechnol. 2014;23(2):345-353. doi:10.1007/s10068-014-0048-x
CrossRef - Dash P, Ghosh G. Amino acid composition, antioxidant and functional properties of protein hydrolysates from Cucurbitaceae seeds. J Food Sci Technol. 2017;54(13):4162-4172. doi:10.1007/s13197-017-2855-6
CrossRef - Je JY, Park SY, Hwang JY, Ahn CB. Amino acid composition and in vitro antioxidant and cytoprotective activity of abalone viscera hydrolysate. J Funct Foods. 2015;16:94-103. doi:10.1016/j.jff.2015.04.023
CrossRef - Esfandi R, Walters ME, Tsopmo A. Antioxidant properties and potential mechanisms of hydrolyzed proteins and peptides from cereals. Heliyon. 2019;5(4):e01538. doi:10.1016/j.heliyon.2019.e01538
CrossRef - Xie J, Du M, Shen M, Wu T, Lin L. Physico-chemical properties, antioxidant activities and angiotensin-I converting enzyme inhibitory of protein hydrolysates from Mung bean (Vigna radiate). Food Chem. 2019;270(235):243-250. doi:10.1016/j.foodchem.2018.07.103
CrossRef - He R, Girgih AT, Malomo SA, Ju X, Aluko RE. Antioxidant activities of enzymatic rapeseed protein hydrolysates and the membrane ultrafiltration fractions. J Funct Foods. 2013;5(1):219-227. doi:10.1016/j.jff.2012.10.008
CrossRef - Zou T Bin, He TP, Li H Bin, Tang HW, Xia EQ. The structure-activity relationship of the antioxidant peptides from natural proteins. Molecules. 2016;21(1):1-14. doi:10.3390/molecules21010072
CrossRef - Maestri E, Marmiroli M, Marmiroli N. Bioactive peptides in plant-derived foodstuffs. J Proteomics. 2016;147(March):140-155. doi:10.1016/j.jprot.2016.03.048
CrossRef - Prados IM, Orellana JM, Marina ML, García MC. Identification of Peptides Potentially Responsible for in Vivo Hypolipidemic Activity of a Hydrolysate from Olive Seeds. J Agric Food Chem. 2020;68(14):4237-4244. doi:10.1021/acs.jafc.0c01280
CrossRef - Lammi C, Aiello G, Boschin G, Arnoldi A. Multifunctional peptides for the prevention of cardiovascular disease: A new concept in the area of bioactive food-derived peptides. J Funct Foods. 2019;55:135-145. doi:10.1016/j.jff.2019.02.016
CrossRef - García MC, Endermann J, González-García E, Marina ML. HPLC-Q-TOF-MS Identification of Antioxidant and Antihypertensive Peptides Recovered from Cherry (Prunus cerasus L.) Subproducts. J Agric Food Chem. 2015;63(5):1514-1520. doi:10.1021/jf505037p
CrossRef - Zheng L, Zhao Y, Dong H, Su G, Zhao M. Structure-activity relationship of antioxidant dipeptides: Dominant role of Tyr, Trp, Cys and Met residues. J Funct Foods. 2016;21:485-496. doi:10.1016/j.jff.2015.12.003
CrossRef - Liao W, Gu L, Zheng Y, et al. Analysis of the quantitative structure-activity relationship of glutathione-derived peptides based on different free radical scavenging systems. Medchemcomm. 2016;7(11):2083-2093. doi:10.1039/c6md00006a
CrossRef - Sun KL, Gao M, Wang YZ, Li XR, Wang P, Wang B. Antioxidant Peptides From Protein Hydrolysate of Marine Red Algae Eucheuma cottonii: Preparation, Identification, and Cytoprotective Mechanisms on H2O2 Oxidative Damaged HUVECs. Front Microbiol. 2022;13. doi:10.3389/fmicb.2022.791248
CrossRef - Gulcin İ, Alwasel SH. Metal Ions, Metal Chelators and Metal Chelating Assay as Antioxidant Method. Processes. 2022;10(1). doi:10.3390/pr10010132
CrossRef - Lin S, Hu X, Li L, et al. Preparation, purification and identification of iron-chelating peptides derived from tilapia (Oreochromis niloticus) skin collagen and characterization of the peptide-iron complexes. LWT. 2021;149. doi:10.1016/j.lwt.2021.111796
CrossRef - Wu H, Liu Z, Zhao Y, Zeng M. Enzymatic preparation and characterization of iron-chelating peptides from anchovy (Engraulis japonicus) muscle protein. Food Research International. 2012;48(2):435-441. doi:10.1016/j.foodres.2012.04.013
CrossRef - Wu W, Li B, Hou H, Zhang H, Zhao X. Identification of iron-chelating peptides from Pacific cod skin gelatin and the possible binding mode. J Funct Foods. 2017;35:418-427. doi:10.1016/j.jff.2017.06.013
CrossRef - Olsen J V., Ong SE, Mann M. Trypsin cleaves exclusively C-terminal to arginine and lysine residues. Molecular and Cellular Proteomics. 2004;3(6):608-614. doi:10.1074/mcp.T400003-MCP200
CrossRef - Tian Q, Fan Y, Hao L, et al. A comprehensive review of calcium and ferrous ions chelating peptides: Preparation, structure and transport pathways. Crit Rev Food Sci Nutr. 2023;63(20):4418-4430. doi:10.1080/10408398.2021.2001786
CrossRef - Zhang Y, Ding X, Li M. Preparation, characterization and in vitro stability of iron-chelating peptides from mung beans. Food Chem. 2021;349. doi:10.1016/j.foodchem.2021.129101
CrossRef - Samaranayaka AGP, Li-Chan ECY. Food-derived peptidic antioxidants: A review of their production, assessment, and potential applications. J Funct Foods. 2011;3(4):229-254. doi:10.1016/j.jff.2011.05.006
CrossRef - Lammi C, Zanoni C, Scigliuolo GM, D’Amato A, Arnoldi A. Lupin peptides lower low-density lipoprotein (LDL) cholesterol through an up-regulation of the LDL receptor/sterol regulatory element binding protein 2 (SREBP2) pathway at HepG2 cell line. J Agric Food Chem. 2014;62(29):7151-7159. doi:10.1021/jf500795b
CrossRef - Lammi C, Zanoni C, Arnoldi A. IAVPGEVA, IAVPTGVA, and LPYP, three peptides from soy glycinin, modulate cholesterol metabolism in HepG2 cells through the activation of the LDLR-SREBP2 pathway. J Funct Foods. 2015;14:469-478. doi:10.1016/j.jff.2015.02.021
CrossRef - Lammi C, Aiello G, Vistoli G, et al. A multidisciplinary investigation on the bioavailability and activity of peptides from lupin protein. J Funct Foods. 2016;24:297-306. doi:10.1016/j.jff.2016.04.017
CrossRef - Mohan A, Udenigwe CC. Towards the design of hypolipidaemic peptides: Deoxycholate binding affinity of hydrophobic peptide aggregates of casein plastein. J Funct Foods. 2015;18:129-136. doi:10.1016/j.jff.2015.06.064
CrossRef - Ngoh YY, Choi SB, Gan CY. The potential roles of Pinto bean (Phaseolus vulgaris cv. Pinto) bioactive peptides in regulating physiological functions: Protease activating, lipase inhibiting and bile acid binding activities. J Funct Foods. 2017;33:67-75. doi:10.1016/j.jff.2017.03.029
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






