
Introduction
Inhibition of pathogenic bacteria by the microbes that naturally exist in our regular diets can be remarkably beneficial to our health considering the pathogens’ biocontrol and disease prevention. Foods that offer such additional benefits than providing mere energy are also highly desirable to the health-conscious consumers.1 When the pathogen-inhibition is of concern, lactic acid bacteria (LAB) comes before all other microbes for their strong antimicrobial activity reported against many microorganisms.2 LAB comprise a large group of heterogeneous bacteria that are generally recognized as safe (GRAS) for our consumption.3 Bacteria in this group are generally Gram-positive, catalase-negative, non-spore forming, and devoid of cytochromes.4 They prefer an initial pH of 6 to 7 for their growth and produce a large amount of lactic acid which is the main end product of their anaerobic carbohydrate fermentation.5 Today, most bacteria used as probiotics in the functional foods and therapeutics, or as starter cultures in the industrial food-processing are the LAB.6 LAB have been isolated from a variety of sources but carbohydrate rich environments such as plants, vegetables, and fruits have been frequently reported as their sources of isolation.7,8 Dairy products and fermented foods and beverages also comprise a very good source of LAB.9 Among the various species identified in fermented foods, Lactobacillus appeared to be the most dominant genus of LAB, followed by Pediococcus in most cases.10 While these bacteria offer a wide range of health benefits11, their antimicrobial activity is recognized as the most important property in both starter culture and probiotic applications.12 The antimicrobial activity of lactic acid bacteria is mainly due to the secretion of various organic acids such as lactic acid, formic acid, and acetic acid which reduces the pH, and is thought to be the principal mechanism of pathogen-inhibition.13 They also cause accumulation of molecules like hydrogen peroxide, carbon dioxide, ethanol, acetaldehyde, bacteriocins and bacteriocin-like substances which also possess antimicrobial activities. In addition to providing bioprotection against pathogenic and spoilage microorganisms, the natural LAB of the fermented foods and beverages also help prolonging their shelf life by the reduction of pH and fermentation of sugars into organic acids.14
Isolation of the naturally existing beneficial lactic acid bacteria from foods and their specific applications is an important endeavor. Unless being isolated and proved to possess specific beneficial properties, however, no microbe can be considered for probiotic application. This study was, therefore, designed to obtain and characterize microbes having antimicrobial activity from foods that are commonly eaten and are also likely to carry beneficial LAB species. Borhani in this regard appeared to be an ideal candidate to us since it is a beverage believed to be highly beneficial for health and digestion. Moreover, it is prepared from ingredients of dairy and plant origins both of which have been known to be good sources of LAB.15 Consequently in the present study, we report the isolation of two borhani–associated LAB species Limosilactobacillus fermentum strain LAB–1 and Levilactobacillus brevis strain LAB–5 and their characterization with regards to their antimicrobial activity. Extent of the antimicrobial effect was assessed against several clinically relevant Gram positive and Gram negative test organisms. Strains of these test microbes are often reported to carry resistance to multiple antibiotics (MDR pathogens). Therefore, their inhibition by the native food-associated LAB species demands special importance. The two LAB species, L. fermentum and L. brevis, were both described in previous studies to offer a few beneficial effects including the inhibitory activity against some pathogenic and spoilage microorganisms.16–18 Few strains of both the LAB groups are already being used as starter cultures, and some are also being considered as potential candidates for probiotic applications in functional foods and medicines.19–23 However, their beneficial properties were mostly found to be strain specific.24 Hence, in order to provide specific insights into the probiotic potential of the two isolates, we performed out a careful assessment of their antimicrobial activity which is recognized as the most important prerequisite for a microbe to be considered for probiotic applications.12,25
Materials and Methods
Isolation, purification and preservation of bacteria
For bacterial isolation, five different borhani samples collected from local shops were mixed and plated on de Man, Rogosa and Sharpe (MRS) agar with or without cysteine-supplementation.15 Colonies with unique morphological appearances were selected and streaked on MRS agar to obtain pure cultures. Glycerol stock of each isolate was prepared and preserved at -20°C and -80°C.26,27 Before use, the isolates were revived in MRS broth and subsequently transferred and grown in fresh medium.
PCR, sequencing and identification
The whole genome sequence of LAB-1 and its annotation and identification have been previously reported.28 For 16S rRNA gene sequencing of LAB-5, its genomic DNA was extracted from a single colony as previously described 29. Fragment of the 16S rRNA gene was amplified by conducting PCR with the universal primers 27F and 1492R, and sequencing was performed as described before.30 The 16S rRNA gene sequence was analyzed using NCBI’s BLAST search program against the GenBank database. The identification was based on the sequence similarity, total score, and number of hits obtained against a particular phylotype.27
Accession numbers
The 16S rRNA gene sequence and WGS can be found in the NCBI database under the accession numbers OM980643 and JAJTII00000000.
Preparation of cell free supernatant (CFS)
CFS of the LAB isolates was prepared by centrifugation of the respective cultures at 10,000×g for 20 min at 4°C. The supernatant so obtained was preserved as 1 mL aliquots at -20°C until used.
Test strains
The Test strains used in antimicrobial assay in the present study are known to have spoilage and/or pathogenic implications and include both Gram-positive and Gram negative microbes listed in Table 1.
Table 1: List of the Test strains used in antimicrobial assays.
| Gram negative |
| Acinetobacter baumannii ATCC 7978 |
| Escherichia coli ATCC 25922 |
| Klebsiella pneumoniae ATCC 13883 |
| Pseudomonas aeruginosa ATCC 9027 |
| Salmonella abony ATCC 14028 |
| Salmonella typhi ATCC 14028 |
| Shigella flexneri ATCC 9199 |
| Gram positive |
| Bacillus cereus ATCC14574 |
| Bacillus subtilis ATCC 6633 |
| Staphylococcus aureus ATCC 6538 |
Agar well diffusion assay
Antimicrobial activity was screened by agar diffusion assay following the protocol reported by Coman et al., 2014 with modifications 31. 1 mL activated culture of each test strain having a cell density equivalent to 0.5 McFarland standard was added to 20 mL of previously autoclaved warm (42- 55°C) Mueller Hinton agar and the suspension was subsequently poured on to petri dishes. Upon solidification of the medium, wells having a diameter of 5 mm was punched using a sterile cork borer and 80 µL of the LAB CFS was placed inside the well. The plates were subjected to a 3 h pre-incubation at 4°C followed by a 24 h incubation at 37° C. Inhibitory activity of the CFS was confirmed by inability of the test strains to grow around the well as indicated by the formation of clear halos. Diameter of the inhibition zones was measured in mm from the edges of the wells.
Broth inhibition assay
Inhibitory effect of the CFS was also assessed by a modified co-incubation broth assay.32 The test strains were activated and grown at 37°C until cell density reached to an OD600 of 1.0. 50 µL of this activated culture was inoculated in 2 mL of nutrient broth containing 400 µL of the CFS and the mixture was incubated at 37°C at 180 rpm in an orbital shaker for 24 h. In the control reaction, sterile MRS broth was used instead of the CFS. Growth of the test strains was monitored by measuring the OD600 at 0 h, 2 h, 4 h, 8 h and 24 h of incubation. Inhibition of growth was determined using the following formula.
Growth inhibition (%) = (AC – AS)/AC x 100, where AC is the absorbance of control and AS is that of sample at 600 nm.
Statistical analysis
All experiments were performed in triplicate and the data are expressed as mean ± standard deviation (SD). The results were statistically evaluated using t-test and P value less than 0.05 was considered to be statistically significant.
Results
Molecular identification and taxonomic assignment
Molecular identification of the LAB-1 strain has been previously reported based on the nucleotide identity of its whole genome sequence and the 16S rRNA gene sequence extracted from the genome sequence 28, whereas identification of the LAB-5 strain was carried out by the amplification and sequencing of a fragment of its 16S rRNA gene followed by sequence identity analysis. Nucleotide BLAST of the 16S rRNA gene sequences of LAB-1 and LAB-5 against the NCBI standard database revealed a high homology, with identity scores more than 99% to the respective genes of several L. fermentum and L brevis strains respectively. The BLAST results with the taxonomic lineage, score, and the number hits obtained for a particular phylotype have been presented in Table 2.
Table 2: Molecular identification of the LAB-1 and LAB-5 strains. Results of nucleotide BLAST of their 16S rRNA gene sequences. Uncultured or unidentified organisms have been removed.
| Isolates | Lineage | Score | No. of Hits |
| LAB-1 | root | 2007 | |
| .Bacteria | 2004 | ||
| ..Bacilli | 1997 | ||
| …Lactobacillaceae | 1995 | ||
| ….Limosilactobacillus | 1985 | ||
| …..Limosilactobacillus fermentum | 2368 | 1973 | |
| …..Limosilactobacillus oris | 2368 | 1 | |
| …..Limosilactobacillus fermentum 3872 | 2368 | 1 | |
| …..Limosilactobacillus fermentum F-6 | 2368 | 1 | |
| …..Limosilactobacillus fermentum CECT 5716 | 2368 | 1 | |
| …..Limosilactobacillus fermentum IFO 3956 | 2368 | 1 | |
| …..Limosilactobacillus sp. | 2303 | 7 | |
| ….Lactobacillus sp. | 2290 | 1 | |
| ….Lacticaseibacillus paracasei | 2287 | 1 | |
| ….Lactobacillus delbrueckii subsp. bulgaricus | 2285 | 1 | |
| ….Lactobacillus sp. T17/4F | 2283 | 1 | |
| ….Lactobacillus sp. JCM 7751 | 2276 | 1 | |
| ….Lactobacillus acidophilus | 2276 | 1 | |
| ….Lactobacillus sp. CR-6AS | 2268 | 1 | |
| LAB-5 | root | 540 | |
| .Bacteria | 539 | ||
| ..Bacilli | 534 | ||
| …Lactobacillaceae | 529 | ||
| ….Levilactobacillus | 509 | ||
| …..Levilactobacillus brevis | 2567 | 502 | |
| …..Levilactobacillus brevis KB290 | 2562 | 1 | |
| …..Levilactobacillus brevis ATCC 367 | 2562 | 1 | |
| …..Levilactobacillus brevis BSO 464 | 2556 | 1 | |
| …..Levilactobacillus sp. | 2556 | 2 | |
| …..Levilactobacillus brevis ATCC 14869 = DSM 20054 | 2556 | 2 | |
| ….Lactobacillus sp. L01 | 2567 | 1 | |
| ….Lactobacillus sp. NBRC 106029 | 2564 | 1 | |
| ….Lactobacillus sp. NBRC 106045 | 2564 | 1 | |
| ….Lactobacillus sp. | 2562 | 8 | |
| ….Lactobacillus sp. SMG65 | 2562 | 1 | |
| ….Lactobacillus sp. NBRC 106022 | 2560 | 1 | |
| ….Lactobacillus sp. NBRC 106046 | 2560 | 1 | |
| ….Lactobacillus sp. ABRIINW.F58 | 2560 | 1 | |
| ….Lactobacillus sp. NBRC 107227 | 2558 | 1 | |
| ….Lactobacillus sp. safa1 | 2556 | 1 | |
| ….Lactobacillus sp. SMG100 | 2556 | 1 | |
| ….Lactobacillus sp. NBRC 106044 | 2556 | 1 | |
| …Bacillus amyloliquefaciens | 2562 | 1 | |
|
|
…Weizmannia coagulans | 2556 |
1 |
Pathogen inhibition in agar diffusion assay
Antimicrobial activity of the two lactic acid bacteria was primarily tested by agar diffusion assay using the cell free supernatant (CFS) against seven Gram-negative (A. baumannii, E. coli, K. pneumoniae, P. aeruginosa, S. abony, S. typhi, S. flexneri) and three Gram-positive bacteria (B. cereus, B. subtilis, S. aureus). Presence of the inhibitory activity was found in the CFS of both isolates with a broad antimicrobial spectrum (Table 3). The L. fermentum strain was able to inhibit all the test strains with a minimum inhibition zone of 9 mm against most of the pathogens. The L. brevis strain showed similar antimicrobial spectrum but with relatively smaller inhibition zones (<6 mm) and a very minor zone against A. baumannii. Both isolates were most effective against P. aeruginosa in the diffusion assay.
Table 3: Inhibition zone (mm) produced against the test strains in agar diffusion assay. The asterisk denotes a statistically significant difference between the two isolates at the 0.05 level.
| Test strains | Inhibition zone (±SD) | |
| LAB-1 | LAB-5 | |
| Gram negative | ||
| A. baumannii | 9 (±1) | Not measured |
| E. coli | 8.67 (±1.16) | 3.34 (±0.58) |
| K. pneumoniae | 9.34 (±1.53) | 3 (±2.65) |
| P. aeruginosa | 10.67 (±1.16) | 5.67 (±0.58) |
| S. abony* | 7.67 (±2.09) | 3.34 (±0.58) |
| S. typhi | 6.34 (±0.58) | 3.67 (±0.58) |
| S. flexneri | 9.5 (±0.5) | 1.34 (±1.16) |
| Gram positive | ||
| B. cereus | 10 (±1.74) | 1 ( ±1.74) |
| B. subtilis | 9.67 (±1.53) | 3 ( ±2.65) |
| S. aureus* | 5 (±1.74) | 3.5 (±1.33) |
Pathogen inhibition in broth assay
To obtain a more precise and more direct evaluation of inhibition, the antimicrobial effect was further estimated by broth inhibition assay in which growth of the above pathogenic microbes was tracked from 0 to 24 h in presence of the CFS. The growth rate and the inhibition kinetics are presented in Figure 1 and 2. The test strains were almost completely suppressed by the L. fermentum CFS after 2, 4, and 8 h of incubation. In a few of the test strains, e.g., E. coli, K. pneumoniae, P. aeruginosa, S. typhi, S. flexneri, and S. aureus, small growths were observed after 24 h which probably was due to the decreasing amount of the antimicrobial substances being used up over time (Figure 1). The relatively less affected test strain was found to be S. aureus followed by S. typhi (Figure 2). Similar to the findings in the agar diffusion assay, antimicrobial activity of L. brevis was found to be less effective than L. fermentum in all cases. The isolate could largely inhibit the pathogens’ growth until 4h or 8h, and a reduced growth was observed afterwards (Figure 1). A. baumannii and S. aureus exhibited relatively better resistance towards the inhibitory activity of L. fermentum (Figure 2).
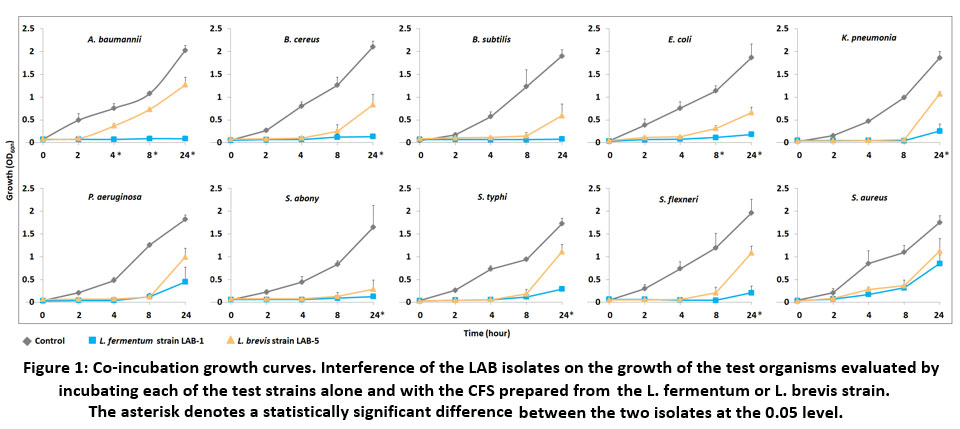 |
Figure 1: Co-incubation growth curves. Interference of the LAB isolates on the growth of the test organisms evaluated by incubating each of the test strains alone and with the CFS prepared from the L. fermentum or L. brevis strain. |
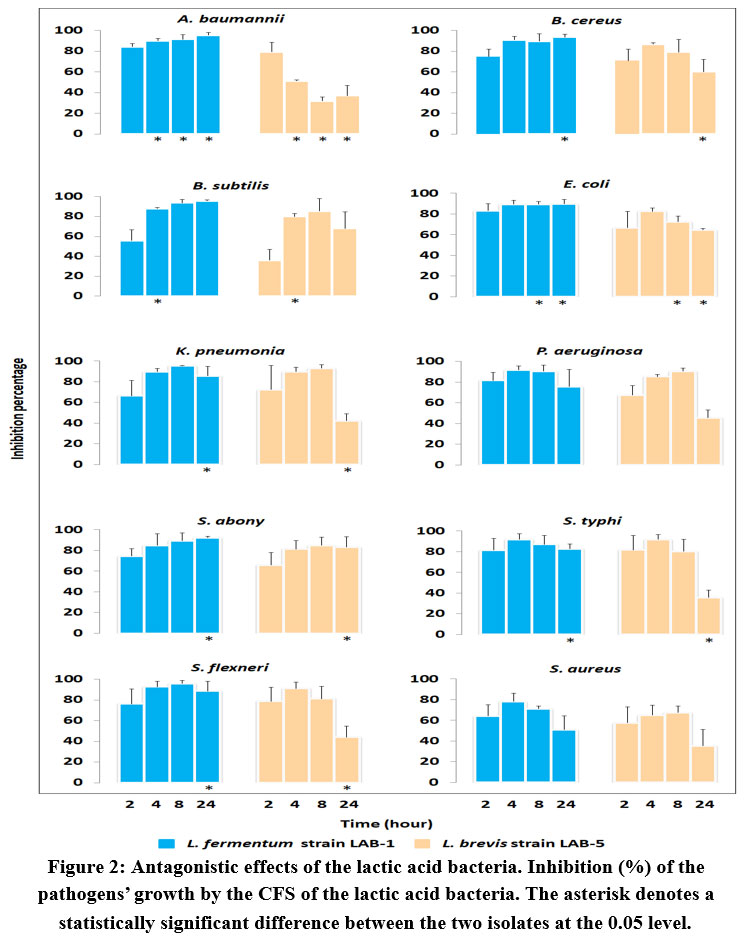 |
Figure 2: Antagonistic effects of the lactic acid bacteria. Inhibition (%) of the pathogens’ growth by the CFS of the lactic acid bacteria. |
Discussion
In the present study, we have assessed antimicrobial activity of two lactic acid bacterial strains, L. fermentum LAB-1 and L. brevis LAB-5 indigenous to the popular South Asian beverage borhani. The two lactic acid bacteria showed a broad spectrum of antimicrobial activity with different extents of inhibitory efficiencies. Both isolates could significantly impede the growth of all the test organisms in both primary and secondary assessments. Although the inhibitory effects varied against specific pathogens in the two methods, the degree of inhibition was generally found same, i.e., the L. fermentum strain appeared to be more efficient than the L. brevis strain in both assay methods. The inhibition area in the agar diffusion assay was at least two to three fold higher with the use of L. fermentum CFS than that obtained with L. brevis (Table 3). The diffusion assay is an easy, rapid and one of the most widely used methods for the assessment of antimicrobial activity in microbial and plant extracts.33 However, this technique relies, to some extent, on smooth diffusion of the sample across the agar and movement of certain antimicrobial compounds is hampered by their interaction with the extrinsic substances or contaminants present in the agar.32 Hence, inhibitory activity of the lactic acid bacteria was further examined by broth inhibition assay in which cell-free culture filtrate of each isolate was co-cultured with the pathogenic microbes and changes in the pathogens’ growth were recorded. The growth stopped almost completely for a significant period of time which confirmed presence of strong antimicrobial activity in the lactic acid bacteria.
Isolation of L. fermentum and L. brevis from dairy products and other sources were previously described. Their antagonistic activity against a few bacterial pathogens has also been demonstrated. One of the L. fermentum strains isolated from the oral mucosa of healthy children inhibited six S. aureus strains including three methicillin-resistant strains.34 Another orally isolated L. fermentum strain, L. f1, was also able to inhibit S. aureus.35 The L. fermentum strain ING8 isolated from Dahi, an Indian traditional fermented milk, could inhibit Listeria monocytogenes, B. cereus, and E. coli.36 Another strain which was obtained from cattle feces, L. fermentum F1, showed antimicrobial activity against S. aureus and E. coli.30 Few antibacterial strains of L. brevis have been reported as well. Streptococcus mutans, a pathogen responsible for dental caries was found to be inhibited by a L. brevis strain that was isolated from the Korean sauerkraut kimchi.38 Inhibitory effect of another L. brevis strain obtained from an Egyptian dairy food was documented against E. coli, S. aureus and S. typhi.17
In the present study, antimicrobial activity of the LAB isolates was examined against a relatively large number of clinically relevant pathogens that are known to cause a range of serious infections in humans. A. baumannii, for example, has been frequently reported in recent years as a causative agent of pneumonia, meningitis, bacteremia, urinary tract infections (UTIs), wound infections, and infections of the skin and soft tissue.39,40 K. pneumoniae, another major pathogen used in the antimicrobial assay, also causes several nosocomial infections and is responsible for pneumonia, UTIs, bloodstream infections and sepsis.41 Moreover, these infections have the ability to metastatically spread and have high rates of morbidity and mortality. Another opportunistic pathogen, P. aeruginosa, is also known to cause serious infections such as malignant external otitis, endophthalmitis, endocarditis, meningitis, pneumonia, and septicemia.42 One of the two Salmonella serovars used in the antimicrobial assays, S. typhi, causes the life-threatening disease typhoid fever.43,44 The other serovar, S. abony, is relatively innocuous; but recently it has been implicated with severe invasive disease and lung abscess.45 Infections with S. flexneri may also develop into life-threatening condition if not carefully treated.46 The three Gram positive pathogens used in this work, B. cereus, B. subtilis, S. aureus, are commonly recognized as food poisoning agents and responsible mainly for bacteremia, gastrointestinal and pulmonary illnesses, and skin infections as well47–50 An additional concern regarding the above infections is the recent advent of multidrug resistance (MDR) in the pathogenic microbes.44,51 All the test pathogens used in the present study for antimicrobial assays are known to carry resistance towards multiple antibiotics.52–61 As a result, treatment of their associated diseases has become very challenging with a few therapeutic options left. In this perspective, natural presence of the antimicrobial lactic acid bacteria in foods and beverages, or their exogenous application in starter culture, functional foods and therapeutics would be of particular benefit to help build protection from the pathogens.
Conclusion
Findings of the present study suggested beneficial lactic acid bacteria to be present in the traditional South Asian beverage borhani. The two species of lactic acid bacteria, Limosilactobacillus fermentum LAB-1 and Levilactobacillus brevis LAB-5 which were isolated from borhani, showed strong antimicrobial activity in agar-diffusion and co-incubation assays. Both isolates could inhibit a wide spectrum of pathogens with a prolonged antimicrobial activity which highlights their important role in enhancing food safety. Their inhibitory effects against spoilage microbes suggest their potential application as food preservatives as well. Upon further in-vivo assessment of their various functional properties, the isolates could be considered as starter culture, adjunct culture or probiotic culture in the fermented foods, functional foods and medications. It is also worth noting that presence of such beneficial lactic acid bacteria as the resident microflora in borhani indicate the potential of getting additional health benefits from this popular beverage.
Acknowledgment
The authors would like to thank Prof. Dr. Nurul Islam, Dept. of Psychology, and Mr. Md. Habibur Rahaman, Dept. of Statistics, University of Chittagong for their help. The authors also thank Nabila Rahman, Subrina Akter Zedny, Mohammad Nazmul Ahmed Chowdhury and all members of the Biochemistry and Pathogenesis of Microbes – BPM Research Group, Bangladesh for their various supports in this project.
Conflicts of Interest
The authors have no conflicts of interest to declare
Funding statement
This project was supported by University of Chittagong, Chattogram 4331, Bangladesh via its Research and Publication Cell to TJH.
References
- Siró I., Kápolna E., Kápolna B., Lugasi A. Functional food. Product development, marketing and consumer acceptance–a review. Appetite. 2008;51(3):456-467. DOI:10.1016/j.appet.2008.05.060
CrossRef - Adeniyi B.A., Adetoye A., Ayeni F.A. Antibacterial activities of lactic acid bacteria isolated from cow faeces against potential enteric pathogens. Afr Health Sci. 2015;15(3):888-895. DOI:10.4314/ahs.v15i3.24
CrossRef - Zacharof M.P., Lovitt R.W. Bacteriocins Produced by Lactic Acid Bacteria a Review Article. APCBEE Procedia. 2012;2:50-56. DOI:10.1016/j.apcbee.2012.06.010
CrossRef - Papadimitriou K., Alegría Á., Bron P.A., Angelis M. de, Gobbetti M., Kleerebezem M., Lemos J.A., Linares D.M., Ross P., Stanton C., Turroni F., Sinderen D. van, Varmanen P., Ventura M., Zúñiga M., Tsakalidou E., Kok J. Stress Physiology of Lactic Acid Bacteria. Microbiol Mol Biol Rev. 2016;80(3):837-890. DOI:10.1128/MMBR.00076-15
CrossRef - Schillinger U., Holzapfel W.H., Björkroth K.J. Lactic acid bacteria. In: Blackburn C de W, ed. Food Spoilage Microorganisms. Woodhead Publishing Series in Food Science, Technology and Nutrition. Woodhead Publishing; 2006:541-578. DOI:10.1533/9781845691417.5.541
CrossRef - Evivie S.E., Huo G.C., Igene J.O., Bian X. Some current applications, limitations and future perspectives of lactic acid bacteria as probiotics. Food Nutri Res. 2017;61(1):1318034. DOI:10.1080/16546628.2017.1318034
CrossRef - Florou-Paneri P., Christaki E., Bonos E. Lactic Acid Bacteria as Source of Functional Ingredients. Lactic Acid Bacteria – R & D for Food, Health and Livestock Purposes. Published online January 30, 2013. DOI:10.5772/47766
CrossRef - Hossain T.J. Functional genomics of the lactic acid bacterium Limosilactobacillus fermentum LAB-1: metabolic, probiotic and biotechnological perspectives. Published online August 6, 2022. Accessed August 6, 2022. https://papers.ssrn.com/abstract=4183480
CrossRef - Ağagündüz D., Yılmaz B., Şahin T.Ö., Güneşliol B.E., Ayten Ş., Russo P., Spano G., Rocha J.M., Bartkiene E., Özogul F. Dairy Lactic Acid Bacteria and Their Potential Function in Dietetics: The Food–Gut-Health Axis. Foods. 2021;10(12):3099. DOI:10.3390/foods10123099
CrossRef - Tamang J.P. Biochemical and modern identification techniques | Microfloras of Fermented Foods. In: Encyclopedia of Food Microbiology. 2nd ed. Academic Press; 2014:250-258. Accessed February 3, 2022. https://www.sciencedirect.com/science/article/pii/B9780123847300000380
CrossRef - Gilliland S.E. Health and nutritional benefits from lactic acid bacteria. FEMS Microbiol Rev. 1990;7(1-2):175-188. DOI:10.1111/j.1574-6968.1990.tb04887.x
CrossRef - Šušković J., Kos B., Beganović J., Leboš Pavunc A., Habjanič K., Matošić S. Antimicrobial activity–the most important property of probiotic and starter lactic acid bacteria. Food Technology and Biotechnology. 2010;48(3):296-307.
CrossRef - Chen C.C., Lai C.C., Huang H.L., Huang W.Y., Toh H.S., Weng T.C., Chuang Y.C., Lu Y.C., Tang H.J. Antimicrobial Activity of Lactobacillus Species Against Carbapenem-Resistant Enterobacteriaceae. Frontiers in Microbiology. 2019;10. Accessed February 3, 2022. https://www.frontiersin.org/article/10.3389/fmicb.2019.00789
- Gänzle M.G. Lactic metabolism revisited: metabolism of lactic acid bacteria in food fermentations and food spoilage. Current Opinion in Food Science. 2015;2:106-117. DOI:10.1016/j.cofs.2015.03.001
CrossRef - Hossain T.J., Nafiz I.H., Mozumder H.A., Ali F., Rahman N., Khan M.S. Antipathogenic action and antibiotic sensitivity pattern of the borhani-associated lactic acid bacterium Weissella confusa LAB-11. Published online July 13, 2022. Accessed September 14, 2022. https://papers.ssrn.com/abstract=4161742
CrossRef - Schurr B.C., Hahne H., Kuster B., Behr J., Vogel R.F. Molecular mechanisms behind the antimicrobial activity of hop iso-α-acids in Lactobacillus brevis. Food Microbiology. 2015;46:553-563. DOI:10.1016/j.fm.2014.09.017
CrossRef - Rushdy A.A., Gomaa E.Z. Antimicrobial compounds produced by probiotic Lactobacillus brevis isolated from dairy products. Ann Microbiol. 2013;63(1):81-90. DOI:10.1007/s13213-012-0447-2
- Naghmouchi K., Belguesmia Y., Bendali F., Spano G., Seal B.S., Drider D. Lactobacillus fermentum: a bacterial species with potential for food preservation and biomedical applications. Critical Reviews in Food Science and Nutrition. 2020;60(20):3387-3399. DOI:10.1080/10408398.2019.1688250
CrossRef - Minj J., Chandra P., Paul C., Sharma R.K. Bio-functional properties of probiotic Lactobacillus: current applications and research perspectives. Critical Reviews in Food Science and Nutrition. 2021;61(13):2207-2224. DOI:10.1080/10408398.2020.1774496
CrossRef - Munekata P.E.S., Pateiro M., Zhang W., Domínguez R., Xing L., Fierro E.M., Lorenzo J.M. Autochthonous Probiotics in Meat Products: Selection, Identification, and Their Use as Starter Culture. Microorganisms. 2020;8(11):1833. DOI:10.3390/microorganisms8111833
CrossRef - Banić M., Uroić K., Pavunc A.L., Novak J., Zorić K., Durgo K., Petković H., Jamnik P., Kazazić S., Kazazić S. Characterization of S-layer proteins of potential probiotic starter culture Lactobacillus brevis SF9B isolated from sauerkraut. LWT. 2018;93:257-267.
CrossRef - Behera S.S., Panda S.K. Ethnic and industrial probiotic foods and beverages: efficacy and acceptance. Current Opinion in Food Science. 2020;32:29-36. DOI:10.1016/j.cofs.2020.01.006
CrossRef - Uroić K., Novak J., Hynönen U., Pietilä T.E., Pavunc A.L., Kant R., Kos B., Palva A., Šušković J. The role of S-layer in adhesive and immunomodulating properties of probiotic starter culture Lactobacillus brevis D6 isolated from artisanal smoked fresh cheese. LWT-Food Science and Technology. 2016;69:623-632.
CrossRef - dos Santos C.I., Campos C.D.L., Nunes-Neto W.R., do Carmo M.S., Nogueira F.A.B., Ferreira R.M., Costa E.P.S., Gonzaga L.F., Araújo J.M.M., Monteiro J.M., Monteiro C.R.A.V., Platner F.S., Figueiredo I.F.S., Holanda R.A., Monteiro S.G., Fernandes E.S., Monteiro A.S., Monteiro-Neto V. Genomic Analysis of Limosilactobacillus fermentum ATCC 23271, a Potential Probiotic Strain with Anti-Candida Activity. Journal of Fungi. 2021;7(10):794. DOI:10.3390/jof7100794
CrossRef - Prabhurajeshwar C., Chandrakanth K. Evaluation of antimicrobial properties and their substances against pathogenic bacteria in-vitro by probiotic Lactobacilli strains isolated from commercial yoghurt. Clinical Nutrition Experimental. 2019;23:97-115. DOI:10.1016/j.yclnex.2018.10.001
CrossRef - Hossain T.J., Alam M.K., Sikdar D. Chemical and microbiological quality assessment of raw and processed liquid market milks of Bangladesh. Continental Journal of Food Science and Technology. 2011;5(2):6-17. DOI:10.5281/zenodo.5568945
- Hossain T.J., Das M., Ali F., Chowdhury S.I., Zedny S.A. Substrate preferences, phylogenetic and biochemical properties of proteolytic bacteria present in the digestive tract of Nile tilapia (Oreochromis niloticus). AIMSMICRO. 2021;7(4):528-545. DOI:10.3934/microbiol.2021032
CrossRef - Hossain T.J. Genome-Sequence, Annotation and Phylogenetic Insights of the Lactic Acid Bacterium Limosilactobacillus Fermentum Strain LAB1 Obtained from the Dairy Beverage Borhani. Social Science Research Network; 2022. Accessed December 30, 2021. http://ssrn.com/abstract=3996663
CrossRef - Hossain T.J., Chowdhury S.I., Mozumder H.A., Chowdhury M.N.A., Ali F., Rahman N., Dey S. Hydrolytic exoenzymes produced by bacteria isolated and identified from the gastrointestinal tract of Bombay duck. Front Microbiol. 2020;11(11:2097). DOI:10.3389/fmicb.2020.02097
CrossRef - Ali F., Sharup Das, Tanim Jabid Hossain, Sumaiya Islam Chowdhury, Subrina Akter Zedny, Tuhin Das, Mohammad Nazmu Ahmed Chowdhury, Mohammad Seraj Uddin. Production Optimization, Stability, and Oil Emulsifying Potential of Biosurfactants from Selected Bacteria Isolated from Oil Contaminated Sites. Royal Society Open Science. 2021;8(211003). DOI:10.1098/rsos.211003
CrossRef - Coman M.M., Verdenelli M.C., Cecchini C., Silvi S., Orpianesi C., Boyko N., Cresci A. In vitro evaluation of antimicrobial activity of Lactobacillus rhamnosus IMC 501(®) , Lactobacillus paracasei IMC 502(®) and SYNBIO(®) against pathogens. J Appl Microbiol. 2014;117(2):518-527. DOI:10.1111/jam.12544
CrossRef - Fredua-Agyeman M., Gaisford S. Assessing inhibitory activity of probiotic culture supernatants against Pseudomonas aeruginosa: a comparative methodology between agar diffusion, broth culture and microcalorimetry. World J Microbiol Biotechnol. 2019;35(3):49. DOI:10.1007/s11274-019-2621-1
CrossRef - Balouiri M., Sadiki M., Ibnsouda S.K. Methods for in vitro evaluating antimicrobial activity: A review. Journal of Pharmaceutical Analysis. 2016;6(2):71-79. DOI:10.1016/j.jpha.2015.11.005
CrossRef - Kang M.S., Lim H.S., Oh J.S., Lim Y. jin, Wuertz-Kozak K., Harro J.M., Shirtliff M.E., Achermann Y. Antimicrobial activity of Lactobacillus salivarius and Lactobacillus fermentum against Staphylococcus aureus. Pathogens and Disease. 2017;75(2):ftx009. DOI:10.1093/femspd/ftx009
CrossRef - Georgieva R., Yocheva L., Tserovska L., Zhelezova G., Stefanova N., Atanasova A., Danguleva A., Ivanova G., Karapetkov N., Rumyan N., Karaivanova E. Antimicrobial activity and antibiotic susceptibility of Lactobacillus and Bifidobacterium spp. intended for use as starter and probiotic cultures. Biotechnology & Biotechnological Equipment. 2015;29(1):84-91. DOI:10.1080/13102818.2014.987450
CrossRef - Pakroo S., Tarrah A., Takur R., Wu M., Corich V., Giacomini A. Limosilactobacillus fermentum ING8, a Potential Multifunctional Non-Starter Strain with Relevant Technological Properties and Antimicrobial Activity. Foods. 2022;11(5):703. DOI:10.3390/foods11050703
CrossRef - Zeng X.Q., Pan D.D., Zhou P.D. Functional Characteristics of Lactobacillusfermentum F1. Curr Microbiol. 2011;62(1):27-31. DOI:10.1007/s00284-010-9669-3
CrossRef - Jang H.J., Kim J.H., Lee N.K., Paik H.D. Inhibitory effects of Lactobacillus brevis KU15153 against Streptococcus mutans KCTC 5316 causing dental caries. Microbial Pathogenesis. 2021;157:104938. DOI:10.1016/j.micpath.2021.104938
CrossRef - Eliopoulos G.M., Maragakis L.L., Perl T.M. Acinetobacter baumannii: epidemiology, antimicrobial resistance, and treatment options. Clinical infectious diseases. 2008;46(8):1254-1263.
CrossRef - Morris F.C., Dexter C., Kostoulias X., Uddin M.I., Peleg A.Y. The Mechanisms of Disease Caused by Acinetobacter baumannii. Frontiers in Microbiology. 2019;10. Accessed March 10, 2022. https://www.frontiersin.org/article/10.3389/fmicb.2019.01601
CrossRef - Bengoechea J.A., Sa Pessoa J. Klebsiella pneumoniae infection biology: living to counteract host defences. FEMS Microbiology Reviews. 2019;43(2):123-144. DOI:10.1093/femsre/fuy043
CrossRef - Lin J.J., Wu C.T., Hsia S.H., Lin K.L., Chiu C.H. Community-acquired Pseudomonas meningitis causes acute obstructive hydrocephalus. Childs Nerv Syst. 2009;25(6):723-725. DOI:10.1007/s00381-009-0836-4
CrossRef - Galán J.E. Typhoid toxin provides a window into typhoid fever and the biology of Salmonella Typhi. Proceedings of the National Academy of Sciences. 2016;113(23):6338-6344.
CrossRef - Ali F., Silvy T.N., Hossain T.J., Uddin M.K., Uddin M.S. Prevalence and antimicrobial resistance phenotypes of Salmonella species recovered at various stages of broiler operations in Hathazari, Bangladesh. Int J One Health. 2021;7(2):158-164. DOI:10.14202/IJOH.2021
CrossRef - Pitiriga V., Dendrinos J., Nikitiadis E., Vrioni G., Tsakris A. First Case of Lung Abscess due to Salmonella enterica Serovar Abony in an Immunocompetent Adult Patient. Case Reports in Infectious Diseases. 2016;2016:e3159031. DOI:10.1155/2016/3159031
CrossRef - Pigott D.C. Foodborne Illness. Emergency Medicine Clinics of North America. 2008;26(2):475-497. DOI:10.1016/j.emc.2008.01.009
CrossRef - Turnbull P.C.B. Bacillus. In: Baron S, ed. Medical Microbiology. 4th ed. University of Texas Medical Branch at Galveston; 1996. Accessed March 11, 2022. http://www.ncbi.nlm.nih.gov/books/NBK7699/
CrossRef - Bottone E.J. Bacillus cereus, a Volatile Human Pathogen. Clinical Microbiology Reviews. 2010;23(2):382-398. DOI:10.1128/CMR.00073-09
CrossRef - Kim H.K., Missiakas D., Schneewind O. Mouse models for infectious diseases caused by Staphylococcus aureus. Journal of Immunological Methods. 2014;410:88-99. DOI:10.1016/j.jim.2014.04.007
CrossRef - Tong S.Y.C., Davis J.S., Eichenberger E., Holland T.L., Fowler V.G. Staphylococcus aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin Microbiol Rev. 2015;28(3):603-661. DOI:10.1128/CMR.00134-14
CrossRef - Klemm E.J., Wong V.K., Dougan G. Emergence of dominant multidrug-resistant bacterial clades: Lessons from history and whole-genome sequencing. Proceedings of the National Academy of Sciences of the United States of America. 2018;115(51):12872. DOI:10.1073/pnas.1717162115
CrossRef - Hernández-González I.L., Castillo-Ramírez S. Antibiotic-resistant Acinetobacter baumannii is a One Health problem. The Lancet Microbe. 2020;1(7):e279. DOI:10.1016/S2666-5247(20)30167-1
CrossRef - Kidd T.J., Mills G., Sá-Pessoa J., Dumigan A., Frank C.G., Insua J.L., Ingram R., Hobley L., Bengoechea J.A. A Klebsiella pneumoniae antibiotic resistance mechanism that subdues host defences and promotes virulence. EMBO Mol Med. 2017;9(4):430-447.
CrossRef - Pang Z., Raudonis R., Glick B.R., Lin T.J., Cheng Z. Antibiotic resistance in Pseudomonas aeruginosa: mechanisms and alternative therapeutic strategies. Biotechnol Adv. 2019;37(1):177-192. DOI:10.1016/j.biotechadv.2018.11.013
CrossRef - Cunha-Neto A. da, Carvalho L.A., Carvalho R.C.T., Dos Prazeres Rodrigues D., Mano S.B., Figueiredo E.E. de S., Conte-Junior C.A. Salmonella isolated from chicken carcasses from a slaughterhouse in the state of Mato Grosso, Brazil: antibiotic resistance profile, serotyping, and characterization by repetitive sequence-based PCR system. Poult Sci. 2018;97(4):1373-1381. DOI:10.3382/ps/pex406
CrossRef - Eng S.K., Pusparajah P., Mutalib N.S.A., Ser H.L., Chan K.G., Lee L.H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Frontiers in Life Science. 2015;8(3):284-293. DOI:10.1080/21553769.2015.1051243
CrossRef - Nguyen T.V., Le P.V., Le C.H., Weintraub A. Antibiotic Resistance in Diarrheagenic Escherichia coli and Shigella Strains Isolated from Children in Hanoi, Vietnam. Antimicrobial Agents and Chemotherapy. 2005;49(2):816-819. DOI:10.1128/AAC.49.2.816-819.2005
CrossRef - Ranjbar R., Farahani A. Shigella: Antibiotic-Resistance Mechanisms And New Horizons For Treatment. Infect Drug Resist. 2019;12:3137-3167. DOI:10.2147/IDR.S219755
CrossRef - Crowe-McAuliffe C., Graf M., Huter P., Takada H., Abdelshahid M., Nováček J., Murina V., Atkinson G.C., Hauryliuk V., Wilson D.N. Structural basis for antibiotic resistance mediated by the Bacillus subtilis ABCF ATPase VmlR. Proceedings of the National Academy of Sciences. 2018;115(36):8978-8983.
CrossRef - Schito G.C. The importance of the development of antibiotic resistance in Staphylococcus aureus. Clinical Microbiology and Infection. 2006;12(s1):3-8. DOI:10.1111/j.1469-0691.2006.01343.x
CrossRef - Foster T.J. Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiology Reviews. 2017;41(3):430-449. DOI:10.1093/femsre/fux007
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








