

Introduction
Oxidative damage refers to the damage of cellular proteins, DNA, and membranes arising from the disturbances in cellular redox potential when elevated levels of reactive oxygen species (ROS) are not counterbalanced.1 These ROS are abundant in the human gut and present an abusive, pro-oxidation environment which may be deleterious towards the gut microbiota.2 Evidence suggests that the intestinal microbiota contributes to the metabolic health of the human host and, when aberrant, to the pathogenesis of various gut and metabolic disorders.3 Oxidative stress is implicated for the pathophysiological of gut diseases such as inflammatory bowel disease and Irritable Bowel Syndromes.4 Combating oxidative stress generated in the gut may be instrumental in upkeeping the gut microbiota and preventing the pathogenesis of various gut-related diseases.
Prebiotic, probiotic, postbiotic, and dietary polyphenols are commonly associated with gut health via their interactions with the gut microbiota.5-7 Prebiotics are non-digestible food ingredients that are naturally occurring or selectively fermented. By causing specific composition changes and/or activity of the gastrointestinal microbiota, the prebiotics promote the host health.8 There are many types of prebiotics, common ones used in food products are inulin, fructooligosaccharides (FOS), and galactooligosaccharides (GOS). Inulin and FOS have a linear chain of fructose with β(2→1) linkage and terminal glucose units.8 Inulin has a DP of up to 60, while the DP of FOS is less than 10.8 GOS consists of two to nine β-(1→2 or 3 or 4 or 6) linked galactose with terminal glucose.9 Probiotics are essentially live microorganisms making up the human gut microflora.10 They help restore the healthy gut microbiota after dysbiosis. The most common probiotic strains present in food and supplements belong to those of the lactic acid bacteria, Lactobacillus, and Bifidobacterium species.10 Coincidentally, they colonize the healthy gut from birth.11 These probiotic strains are thought to alleviate different diseases via competitive inhibition of other microbes12, improvement of mucosal barrier function12, and interaction with dendritic cells13. Postbiotic is defined as dead microorganisms and their components that can confer a health benefit on the host.5 It is the soluble products or metabolic byproducts secreted by live bacteria or released after bacterial lysis, usually enzymes, bacterial peptides and proteins, polysaccharides, and organic acids.5 Short-chain fatty acids (SCFAs), such as acetic, propionic, and butyric acids, are organic acids commonly found in postbiotics.14 A total of fifty to one hundred mmol L−1 of these three saturated fatty acids are produced by the healthy gut microbiota daily.15 Dietary polyphenols are phytochemicals naturally present in plants. They are shown to interact extensively with the gut microbiota. Recent studies demonstrate that polyphenols improve gut health by increasing the relative abundance of lactic acid bacteria, such as bifidobacteria and lactobacilli.16 The colon microbiota, in turn, transform the dietary polyphenols into absorbable, bioactive organic acids .16
To date, the effects of prebiotics, probiotics, postbiotics, and dietary polyphenols on the oxidative environment in the gut are less studied interdependently. These bioactives can be present together in the gut and may interact to modulate the oxidative potential in the gut environment. The study evaluated the influence of common dietary polyphenols (quercetin), prebiotics (GOS), probiotics (Lactobacillus acidophilus, LBA), and/ or postbiotics (inanimate LBA) on the cellular oxidative status in the human gut, using the established CACO-2 intestinal epithelial cell model. The same study permuted the combination of polyphenol, prebiotic, probiotic, and postbiotic to evaluate their individual and synergistic cellular antioxidant effects.
Material and Methods
Chemicals and materials
CACO-2 HTB-37™ cells and Lactobacillus acidophilus ATCC® 4356™ (LBA) were acquired from American Type Culture Collection (Manassas, VA, USA). Arachidonic acid (AA), F2-isoprostanes (F2IP), and F2-isoprostanes-d4 were purchased from Cayman Chemical (Ann Arbor, MI, USA). 1,1-diphenyl-2-picrylhydrazyl radical (DPPH), 2,2′-azinobis-(3-ethylbenzthiazoline-6-sulfonate) (ABTS), 2,2′-azobis-(2-amidinopropane) hydrochloride (ABAP), 2,2′-azobis(2-methylpropionamidine) dihydrochloride (AAPH), 2,3,4,5,6-pentafluorophenylbromide, 4-methylpentanoic acid, 12-myristate 13-acetate (PMA), β-nicotinamide adenine dinucleotide (NADH), acetic acid, ammonium ferrous sulfate, bis(trimethylsilyl) trifluoroacetamide, butyric acid, cytochrome c, dimethyl sulfoxide (DMSO), decylamine, Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F12), fetal calf serum (FCS), fluorescein sodium salt, GOS, glutamine, guaiacol, hydrogens peroxide (50% by volume), isooctane, phorbol, MRS medium, MRS agar, penicillin, phosphate-buffered saline (PBS), propionic acid, pyridine, R-phycoerythrin, H3PO4, quercetin (Q), NaH2PO4, sodium orthophosphate, streptomycin, superoxide dimutase (SOD), toluene, trichloroacetic acid (TCA), Tris (2-carboxyethyl)-phosphine hydrochloride (TCEP-HCl), trolox, trypan blue, Tween-20, Vitamin C, and xylenol orange were purchased from Sigma-Aldrich (St. Louis, MO, USA). Acetonitrile, ethyl acetate, methanol, ethanol, and sulfuric acid were purchased from Tedia (Fairfield, OH, USA).
Culture and Inactivation of Lactobacillus acidophilus
LBA was cultured from an initial concentration of 104 CFU/ mL in MRS medium at 5% CO2 at 37 °C for 24 hours. The primary culture was subcultured into the same media for 36 hours at 37 °C with shaking motion (100 rev/min). Viable cell concentrations were measured by spread plate counting in duplicate on MRS agar after incubation at 5% CO2 at 37 °C for 48 hours.
The LBA culture was lyzed by sonication and centrifugation at 2000xg to obtain the LBA postbiotic (iLBA) supernatant.
Culture and treatment of CACO-2 cells
CACO-2 cells were cultured in DMEM/F12 supplemented with heat-inactivated FCS, glutamine, penicillin, and streptomycin to final concentrations of 10%, 2 mmol L-1, 100 U mL-1, and 100 lg mL-1, respectively. The viability of the CACO-2 cells was measured using the lactate dehydrogenase assay17. Cell viability of >98% was used for the in vitro experiments.
Either Q (final concentration, 1 µmol L-1), GOS (final concentration, 4 mg mL-1), LBA (final concentration, 2 x 106 CFU mL-1), iLBA (final concentration, 2 x 106 CFU mL-1), Q+GOS, Q+LBA, Q+iLBA, GOS+LBA, GOS+iLBA, LBA+iLBA, Q+GOS+LBA, Q+GOS+iLBA, Q+LBA+iLBA, GOS+LBA+iLBA or ALL (all at their individual final concentrations) was added into the CACO-2 cells (5×106 cells mL-1 in PBS, 1 mL). The cell mixture was incubated with AA (final concentration, 10 mmol L-1) and AAPH (final concentration, 5 mmol/ L) at 37 °C and 5% carbon dioxide for 0, 1, 2, 5, 12, and 24 hours. At the end of incubation, CACO-2 cell pellets were obtained after centrifugation at 2000xg for 5 min at 4 ºC, washed once with PBS before being lyzed in buffer (30 mM NaH2PO4, adjusted to pH 3.0 with H3PO4) by sonication. The cell lysates were stored at -80 ºC before subsequent analyses.
Antioxidant capacity
The antioxidant capacities of the text mixture on CACO-2 cells were determined using DPPH radical scavenging activity18, Oxygen Radical Absorbance Capacity (ORAC)19, Total Peroxyl Trapping Potential (TRAP)20, and Trolox Equivalent Antioxidant Capacity (TEAC)21.
Briefly, for DPPH radical scavenging assay18, the cell supernatant (1 mL) was added into the freshly prepared ethanolic DPPH solution (48 mg L-1, 9 mL). The change in absorbance at 517 nm was measured. A calibration curve was prepared using vitamin C (0-500 mmol L-1) as the radical scavenger . Absorbance was corrected for the solvent. The DPPH radical scavenging results were expressed in mmol L-1 vitamin C equivalents.
Briefly, for ORAC assay19, cell supernatant (20 µL) and fluorescein sodium salt (200 µL, 81.6 nmol L-1) were added into a well of a 96-well plate. Fluorescence (kexcitation = 485 nm, kemission = 528 nm) was measured in a multi detection microplate reader (Powerscan, Dainippon Sumitomo Pharma, Osaka, Japan). AAPH (75 µL, 200 mM) was then added to the well. Fluorescence was recorded every min over 40 min. A calibration curve was prepared with trolox (0-50 µmol L-1). Fluorescence was corrected for the solvent. The ORAC results were expressed as µmol L-1 trolox equivalents.
Briefly, for TRAP assay20, the cell supernatant (1 mL) was incubated with TRAP reaction mixture (0.43 mg L-1 R-phycoerythrin and 0.3 g L-1 AAPH in 50 mmol L-1 PBS (pH 7.4), 2 mL) at 37 ºC for 5 min. Fluorescence (kexcitation = 540 nm, kemission = 565 nm) was measured in a quartz cuvette. A calibration curve was prepared with vitamin C (0 – 100 mmol L-1). Fluorescence was corrected for the solvent. The TRAP results were expressed as mmol L-1 vitamin C equivalents.
Briefly, for TEAC assay21, the cell supernatant (40 µL) was mixed with the freshly prepared radical solution (2.5 mmol L-1 ABAP and 20 mmol L-1 ABTS stock solution in PBS, 1960 µL). The decrease in absorption at 734 nm was measured over 6 min. A calibration curve was prepared with trolox (0-1 mmol L-1). Absorbance values were corrected for the solvent. The TEAC results were expressed in mmol L-1 trolox equivalents.
All experiments were performed at least in quintuplicate.
Antioxidative enzyme activity
The antioxidant enzyme activity of the treated CACO-2 cells was assessed by determining the enzyme activities of SOD and peroxidases. The CACO-2 cell pellet obtained after the designated treatment was lyzed by sonication at 4 ºC for 5 min. The SOD activity in the lyzed cell supernatant was determined using a commercial SOD colorimetric activity assay kit (Thermo Fisher Scientific, Waltham, MA, USA). The peroxidase activity in the lyzed cells was measured using a commercial peroxidase activity assay kit (Merck, Darmstadt, GERMANY). The SOD and peroxidase enzyme activities were expressed as the percentage of inhibition relative to the untreated control.
Cellular oxidative damage
The amounts of cellular oxidative damage were determined by measuring the inhibition of F2IP, and lipid hydroperoxides (LPO) productions from CACO-2 cells. The cells were incubated with the respective treatments and AA (final concentration, 10 mmol/ L). Untreated control experiments were carried out by incubating cells with AA (final concentration, 10 mmol/ L) only. At the end of the incubation, the cell supernatant and cell lysate were collected and stored at -80 °C before F2IP and LPO analyses. All experiments were performed at least in quintuplicate. F2IP was quantified using stable isotope-labeled Gas Chromatography-Mass Spectrometry.22 The formation of LPO was quantitated using the Ferrous Oxidation-Xylenol Orange assay.23 The amount of cellular oxidative damage was expressed as the percentage of cellular F2IP and LPO production relative to the positive controls.
Quercetin and short-chain fatty acids
The concentrations of SCFAs in each treatment mixture were determined by using gas chromatography according to Chang et al.24 with slight modifications. The treatment mixture (500 µL) was added with 4-methylpentanoic acid (internal standard, 5 mmol L-1, 500 µL), and 1 µL of the sample was injected into a gas chromatograph (GC-7820, Agilent Technologies, USA). The GC was equipped with a flame ionization detector. The SCFAs were separated using a DB-FFAP 122-3232 fused-silica capillary column (Agilent J&W, 30 m × 0.25 µm × 0.25 µm) set at 100 ºC, increased to 180 ºC with 10 ºC/ min. Temperatures for the detector, inlet, and oven were set at 250 ºC, 230 ºC, and 230 ºC, respectively. Nitrogen gas was used as the carrier gas with a flow of 35 mL/ min, while methanol was used as the solvent. Acetic acid (0-10 mmol L-1), propionic acid (0-10 mmol L-1), butyric acid (0-10 mmol L-1), and 4-methylpentanoic acid (internal standard, 5 mmol L-1) were used as standards to identify the peaks and to plot the calibration curves.
Intracellular concentrations of Q and SCFAs were determined in the cell lysates, while the extracellular concentrations were determined in the cell supernatants. Briefly, for quercetin measurement23, the components (500 µL) were separated by reverse-phase high-performance liquid chromatography using a LiChrospher 100 column (RP-18, 5 µm, 4.0 x 250 mm, Agilent Technologies, CA, USA) with sodium orthophosphate (30 mM; adjusted to pH 3 with phosphoric acid): acetonitrile (15:85, v/v) mobile phase (solvent A) with increasing gradient of acetonitrile (solvent B) at a flow rate of 0.8 mL/ min over 20 min (from 0% to 50% B in A). Dual-wavelength detection at 370 and 270 nm was used to detect the B-ring and C-ring contained within the 2-Phenyl-4H-1-benzopyran-4-one structure of quercetin, respectively. The SCFAs were measured using the GC method as previously described. The cell analyte sample (500 µL) was added with 4-methylpentanoic acid as an internal standard (5 mmol L-1, 500 µL), and 1 µL of the sample was injected into the GC. Acetic, propionic, and butyric acids were determined using their respective calibration curves. Total SCFA was presented as the sum of the three measured acids.
Statistical analysis
Statistical analyses were performed using IBM SPSS Statistics version 26.0 (USA). Data were presented as mean ± standard deviation (SD). Differences between more than two groups were compared using ANOVA with the Bonferroni post-hoc test. Correlations between antioxidant activities, Q and SCFA concentrations were determined using Spearman correlation analysis. A significant difference or correlation was observed when p<0.05.
Results
Non-enzymatic antioxidant capacity
Treatments without Q and iLBA did not alter all measured total antioxidant capacities of the cell mixtures compared to the control treatment (Figure 1). Incubation with Q or/ and iLBA significantly augmented the antioxidant capacities, measured as DPPH, ORAC, TEAC, and TRAP, of the CACO-2 cell mixtures compared to those in the absence of Q or/ and iLBA (Figure 1). The strengths of DPPH radical scavenging activity and TEAC over the 24 hour incubation period increased significantly in the order: GOS ≈ LBA ≈ GOS+LBA (treatments without Q and iLBA) < iLBA ≈ GOS+iLBA ≈ LBA+iLBA ≈ GOS+LBA+iLBA (treatments with iLBA in the absence of Q) < Q ≈ Q+GOS ≈ Q+LBA ≈ Q+GOS+LBA (treatments with Q in the absence of iLBA) < Q+iLBA ≈ Q+GOS+iLBA ≈ Q+LBA+iLBA ≈ Q+GOS+LBA+iLBA (treatments with Q and iLBA) (Figures 1a, Supplementary Data Table 1). The ORAC and TRAP strengths over the 24-hour incubation period increased significantly in the order: GOS ≈ LBA ≈ GOS+LBA (treatments without Q and iLBA) < Q ≈ iLBA ≈ Q+GOS ≈ Q+LBA ≈ GOS+iLBA ≈ LBA+iLBA ≈ Q+GOS+LBA ≈ GOS+LBA+iLBA (treatments with Q or iLBA) < Q+iLBA ≈ Q+GOS+iLBA ≈ Q+LBA+iLBA ≈ Q+GOS+LBA+iLBA (treatments with Q and iLBA) (Figures 1, Supplementary Data Table 1). All the measured antioxidant capacities of the Q- and/ or iLBA-treated cell mixtures decreased steadily over the 24-hour incubation (Figure 1). Q- or/ and iLBA-treated cells demonstrated significantly higher total antioxidant capacities than those not treated with Q or iLBA even after the 24-hour incubation period (Figure 1).
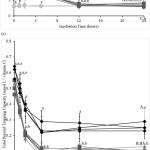 |
Figure 1: (a) 1,1-diphenyl-2-picrylhydrazyl radical scavenging activity (mmol L-1 vitamin C), (b) Oxygen Radical Absorbance Capacity (µmol L-1 trolox), (c) Total Peroxyl Trapping Potential (mmol L-1 vitamin C), and (d) |
Antioxidant enzyme activity
Treatments containing Q increased the SOD and PO enzyme activities of CACO-2 cells that peaked at around 1 hour before gradually lowering to constant activity levels over 24 hours (Figures 2a and 2b). Other treatments did not significantly influence the cellular SOD and PO enzyme activity (Figure 2). The SOD and PO enzyme activities of CACO-2 cells incubated with Q over the 24 hours were significantly higher compared to those not treated with Q (Figures 2a and 2b). GOS, LBA, and iLBA did not influence the SOD and PO enzyme activities in the presence of Q (Figures 2a and 2b, Supplementary Data Table 2).
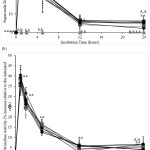 |
Figure 2: (a) Superoxide dismutase enzyme activity and (b) peroxidase enzyme activity (% relative to untreated cells) after incubating CACO-2 cells (5 x 106 cells mL-1) without quercetin and inactivated Lactobacillus acidophilus (●) |
Cellular oxidative damage
The amounts of F2IP and LPO produced by the CACO-2 cells increased over the 24-hour incubation (data not shown). Treatments with Q or/and iLBA significantly inhibited cellular F2IP production compared to those in their absence over the 24-hour incubation period (Figure 3a). The inhibition of F2IP formation was significant between treatments in the order: Untreated ≈ GOS ≈ LBA ≈ GOS+LBA (treatments without Q and iLBA) < iLBA ≈ GOS+iLBA ≈ LBA+iLBA ≈ GOS+LBA+iLBA (treatments with iLBA in the absence of Q) < Q ≈ Q+GOS ≈ Q+LBA ≈ Q+GOS+LBA (treatments with Q in the absence of iLBA) < Q+GOS ≈ iLBA ≈ Q+LBA+iLBA ≈ Q+GOS+LBA+iLBA (treatments with Q and iLBA) (Figure 3a, Supplementary Table 3). Most of the F2IP inhibitory activity occurred in the first 12 hours of the incubation period, with negligible amounts of activity after this period (Figure 3a). F2IP inhibitory activity peaked after 2 hours before gradually decreasing to significantly lower levels after 12-hours (Figure 3a). The F2IP inhibitory activity with Q and iLBA treatments remained significantly higher after 12 and 24 hours of incubation than the other treatments (Figure 3a).
The cellular LPO formations were significantly decreased with treatments involving Q or/and iLBA when compared to those in their absence (Figure 3b). The inhibition of LPO formation by each treatment significantly increased in the order: Untreated ≈ GOS ≈ LBA ≈ GOS+LBA (treatments without Q and iLBA) < iLBA ≈ GOS+iLBA ≈ LBA+iLBA ≈ GOS+LBA+iLBA (treatments with iLBA in the absence of Q) < Q ≈ Q+GOS ≈ Q+LBA ≈ Q+GOS+LBA (treatments with Q in the absence of iLBA) < Q+GOS ≈ iLBA ≈ Q+LBA+iLBA ≈ Q+GOS+LBA+iLBA (treatments with Q and iLBA) (Figure 3b, Supplementary Table 3). LPO formation inhibitions of Q or/ and iLBA treatments peaked between the second and fifth hour (Figure 3b). Treatments with Q or/and iLBA significantly reduced cellular LPO formations at most incubation time points (t=1h to t=12h) compared to the other treatments (Figure 3b). LPO formation inhibitory activity remained significantly higher after 24 hours only for treatments involving both Q and iLBA (Figure 3b).
 |
Figure 3: Amounts of (a) F2-isoprostanes and (b) lipid hydroperoxides (% relative to the positive controls) formed after incubating CACO-2 cells (5 x 106 cells mL-1) without quercetin and inactivated Lactobacillus acidophilus (●) |
Quercetin and short-chain fatty acids
Acetic (4.16±0.26 mmol L-1), propionic (1.61±0.33 mmol L-1), and butyric (1.59±0.16 mmol L-1) acids were present only in iLBA-containing treatment mixtures. The measured SCFA concentrations did not differ significantly between these mixtures.
Only cells incubated with Q contained significant amounts of intracellular Q over the 24-hour incubation period (Figure 4a). Concentrations of intracellular Q increased during the first 2 hours of incubation before decreasing to baseline levels within 12 hours (Figure 4a). The presence of GOS, LBA, and/or iLBA with Q did not influence the intracellular Q concentrations (Figure 4a, Supplementary Data Table 4). The concentrations of Q remained significantly higher within cells incubated with Q than those without Q at the 24th-hour incubation time point (Figure 4a, Supplementary Data Table 4).
The intracellular total SCFA concentrations of cells treated with Q and/ or iLBA were significantly higher than those of the untreated control cells over the 24 incubation hours (Figure 4b). Treatments without Q or iLBA did not significantly influence the intracellular total SCFA concentrations compared to the untreated control cells over the 24 incubation hours (Figure 4b). For cells treated with iLBA, the intracellular total SCFA concentrations peaked around the second-hour mark before declining to steady concentrations towards the end of treatment. The observed steady concentrations remained significantly higher than the untreated control cells at the 24th-hour time point. For the cells incubated with Q in the absence of iLBA, the measured SCFAs peaked at the 5th hour (Figure 4b). The concentrations of intracellular total SCFA within the cells increased in the order: Untreated ≈ GOS ≈ LBA ≈ GOS+LBA (treatments without Q and iLBA) < Q ≈ Q+GOS < iLBA ≈ Q+LBA ≈ LBA+iLBA ≈ GOS+iLBA ≈ Q+GOS+LBA ≈ GOS+LBA+iLBA (treatments with either Q or iLBA) < Q+iLBA ≈ Q+GOS+iLBA ≈ Q+LBA+iLBA ≈ Q+GOS+LBA+iLBA (treatments with Q and iLBA) (Figure 4b, Supplementary Table 4).
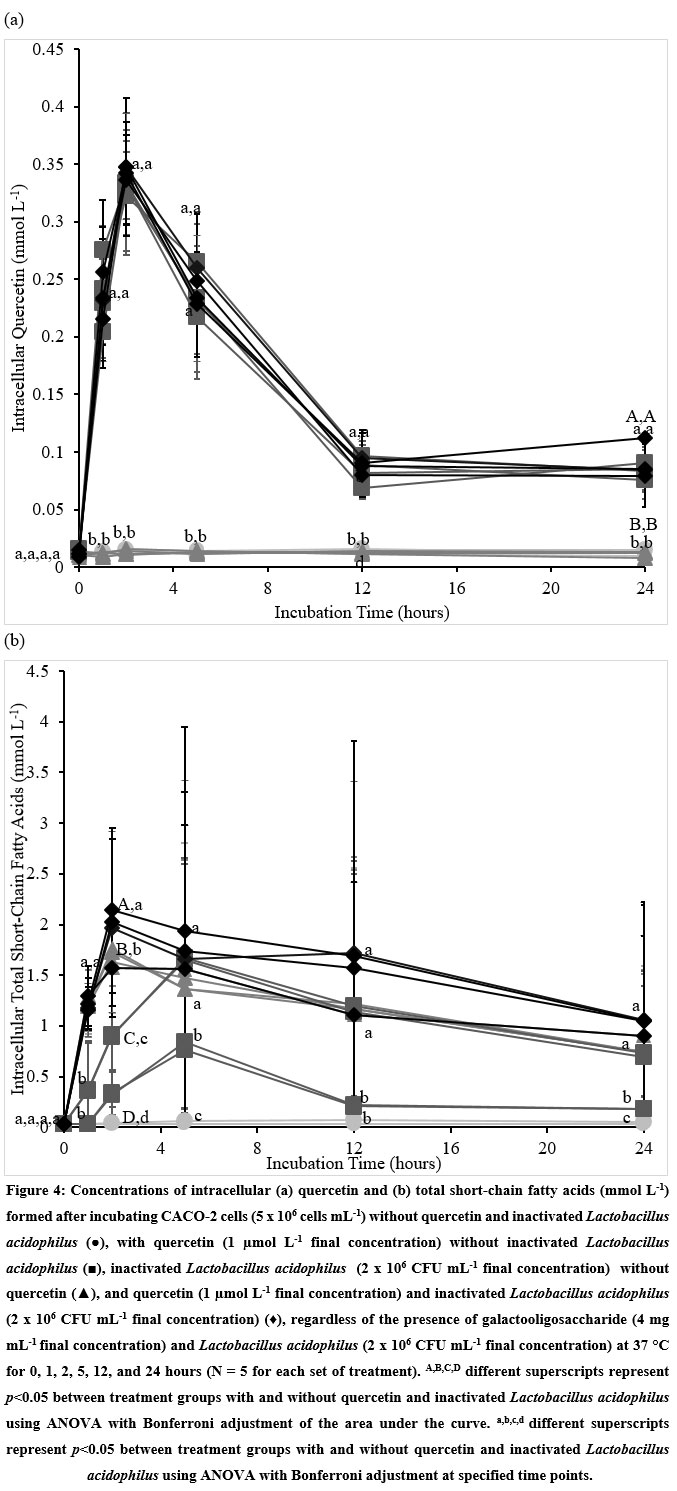 |
Figure 4: Concentrations of intracellular (a) quercetin and (b) total short-chain fatty acids (mmol L-1) formed after incubating CACO-2 cells (5 x 106 cells mL-1) without quercetin and inactivated Lactobacillus acidophilus (●) |
Correlations
Cellular F2IP and LPO inhibitions were significantly correlated with intracellular Q (F2IP, R=0.51, p=0.04; LPO, R=0.62, p=0.03) and propionic acid (F2IP, R=0.42, p=0.03; LPO, R=0.55, p=0.04) concentrations, but not with intracellular acetic acid (F2IP, R=0.32, p=0.23; LPO, R=0.23, p=0.33) concentrations. The measured antioxidant capacities, DPPH, ORAC, TEAC, and TRAP, were positively correlated to intracellular Q (DPPH, R=0.574, p=0.03; ORAC, R=0.327, p=0.04; TEAC, R=0.639, p=0.04; TRAP, R=0.475, p=0.05), propionic acid (DPPH, R=0.435, p=0.04; ORAC, R=0.686, p=0.04; TEAC, R=0.436, p=0.04; R=0.694, p=0.03; TRAP, R=0.578, p=0.04), and butyric acid (DPPH, R=0.656, p=0.04; ORAC, R=0.435, p=0.03; TEAC, R=0.677, p=0.04; R=0.554, p=0.03; TRAP, R=0.435, p=0.03) concentrations.
Discussion
The digestive tract is constantly subjected to oxidative insults from exogenous and endogenous oxidants.2, 25 It is important to maintain oxidative balance within the digestive tract to maintain a healthy microbiota. Similar to in vivo settings, the CACO-2 cells in the experiments were experiencing persistent oxidative damage, as evidenced by the steadily increasing concentrations of F2IP and LPO formed in the cell mixtures over the incubation period. The attenuation of F2IP and LPO formation by treatments with either Q or iLBA suggested that Q and iLBA exerted significant antioxidant activities by protecting the CACO-2 cells from oxidative damages. This was supported by the significant positive correlations between the measured Q concentrations and antioxidant capacities – DPPH, ORAC, TEAC, TRAP, cellular F2IP, and LPO inhibitions. The results from the radical scavenging and antioxidant enzyme activity assays suggest that Q protected the CACO-2 cells from oxidative damage by scavenging free radicals directly and upregulating the cellular antioxidant enzymes, such as SOD and PO, whereas iLBA did so only via scavenging radicals. The four radical scavenging assays used in the experiments differ in their reaction mechanisms and can be employed to evaluate the radical scavenging mechanisms.26 ORAC and TRAP involve the transfer of hydrogen atoms from the probe to the radical, and they measure the radical scavenging capacity via the hydrogen atom transfer (HAT) mechanism.26 DPPH and TEAC, on the other hand, involve the electron transfer between the probe and radicals and thereby determine the radical scavenging activity via the electron transfer (ET) mechanism.26 Radical scavenging by Q and iLBA occurs through HAT and ET mechanisms. Q, like other polyphenols, is a powerful antioxidant in vitro. Q scavenges free radicals through both HAT and ET mechanisms at different rates.27 The DPPH, ORAC, TEAC, and TRAP results of Q agree with the previously computed observations27 that Q is capable of donating H atoms and electrons to free radicals. Q was shown to be a stronger electron donor than iLBA, and both were equally strong as H atom donors. In addition, Q, not iLBA, was shown to augment the activity levels of SOD and PO in the CACO-2 cells. Q elevated SOD activity in alveolar epithelial A54928 and liver Hep2G29 cells. While Q has been studied in numerous disease settings involving oxidative stress, its biological activity is less examined in the human digestive tract. Q significantly inhibited lipopolysaccharide-induced jejunal oxidative stress in broiler chickens by upregulating SOD and glutathione peroxidase levels.30 Q exhibited greater reactivity (forty folds) towards peroxyl radicals in the aqueous solution of neutral pH 7.4 than in the aqueous solution of acidic pH 3.0.31 Accordingly, Q may operate effectively as H and electron donors in the native pH environment of the human small intestine. The reported beneficial effects of dietary polyphenols on gut microbiota32 may be attributed to their antioxidant properties.
The iLBA contained significantly higher concentrations of total SCFA compared to the GOS, LBA, and Q. These SCFAs were not significantly present in the LBA before the inactivation process, and were also absent in the other treatment mixtures. Their presence may account for the observed antioxidant property by iLBA. This deduction is also supported by the observed significant association of propionic and butyric acid concentrations with the cellular oxidative damages and antioxidant capacities. SCFAs inhibited lipopolysaccharide-induced oxidative stress in mesangial cells.33 SCFAs significantly and differentially decreased Nlrp3 inflammasome activation and formation in endothelial cells.34 Organic acid production from postbiotics was positively correlated with the antioxidant activity of the postbiotics derived from Lactiplantibacillus plantarum strains.24 Our results add to the limited data on the potential antioxidant activity of postbiotic SCFAs. Q exhibited comparatively stronger radical scavenging activity and thereby protected the CACO-2 cells to significantly greater extents from oxidative damages than the iLBA. The resonance-stabilized polyphenolic structure makes Q a stronger H atom and electron donor than the aliphatic carboxylic structure of the SCFA, possibly explaining the significant difference in antioxidant strength between Q and iLBA. The ORAC and TRAP results suggest that Q and iLBA are comparable H donors. Q demonstrated to be a stronger electron donor compared to iLBA, as observed by the differences in their DPPH and TEAC results. The absence of effects on the cellular antioxidant enzyme activity by iLBA may also diminish its antioxidant capacity when compared to Q. The difference in significant correlation between the two measured SCFA concentrations and cellular antioxidant capacities and oxidative damage markers suggest that two SCFAs (propionic and butyric acids) exhibited differential antioxidant capacities. More studies should be carried out to evaluate the antioxidant capacity of these SCFAs on the representative CACO-2 cell model. The presence of Q and iLBA presented summative antioxidant protection compared to the presence of either one. They appeared to contribute independently due to the difference in their antioxidant mechanisms. Additional studies are required to elucidate the potential interactions between these molecules in modulating intestinal oxidative status.
Q and SCFAs were shown to accumulate into the cells within the first two hours of incubation, and this accumulation translated to augmented protection against oxidative damages. The antioxidant protection was steadily reduced as the amounts of intracellular Q and SCFAs dissipated over the next 24 hours. The antioxidant capacity remained significantly higher compared to the cells not treated with Q and/ or iLBA after the 24-hour incubation. Q has been demonstrated to be metabolized to simpler organic and phenolic compounds.35 This is evident from the increased concentrations of SCFAs in the Q-treated CACO-2 cells. The phenolic compounds derived from Q metabolism were shown to exhibit antioxidant properties and protected CACO-2 cells against oxidative damage.36 Together, they may explain the heightened antioxidant capacity even after 24 hours.
GOS, used in the study, did not affect the SCFA concentrations in the CACO-2 cell culture within the 24-hour treatment period. Prebiotics have been shown to influence production of SCFAs by pure cultures in vitro.37 Gut microbiota fermented the prebiotics into short-chain fatty acids, including lactic acid, butyric acid, and propionic acid.14 It becomes possible that prebiotics can exert antioxidant effects via the production of antioxidants like SCFAs by the fermentative action of a probiotic. The short treatment duration in this study did not allow significant fermentation of the prebiotic. More studies are required to ascertain the antioxidant effect, whether direct or indirect, of prebiotics, such as GOS. In the study, the effects of oxidative stress by the polyphenol, prebiotic, probiotic, and postbiotic on CACO-2 cells was tested in four ways: (1) direct quantitation of non-enzymatic radical scavenging capacities via DPPH, ORAC, TEAC, and TRAP assays; (2) direct measurement of specific antioxidant enzyme activities via SOD and PO enzymes; (3) measurement of the resulting damage to biomolecules (arachidonic acid and lipid); and (4) measurement of antioxidant levels (Q and total SCFAs). Future clinical or in vivo studies should be conducted to evaluate these in vitro findings. The study results are also limited by the choice of polyphenol (Q), prebiotic (GOS), probiotic (LBA), and postbiotic (iLBA). More future studies are required to evaluate other possible candidates of these families of functional ingredients.
Polyphenol, probiotic, and postbiotic, individually or cumulatively, influence the oxidative status of intestinal epithelial CACO-2 cells and protect them from oxidative damage. The study provides in vitro evidence to support the inclusion of all these functional ingredients to maintain the intestinal oxidative status and improve gut microbiota, and henceforth digestive health.
Acknowledgment
None
Conflict of Interest
The authors have no conflict of interest.
Funding Source
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
- Cassidy L., Fernandez F., Johnson J. B., Naiker M., Owoola A. G., Broszczak D. A. Oxidative Stress in Alzheimer’s Disease: A Review on Emergent Natural Polyphenolic Therapeutics. Complement Therap Med. 2020;49:102294-102300. https://doi.org/10.1016/j.ctim.2019.102294
CrossRef - Bhattacharyya A., Chattopadhyay R., Mitra S., Sheila E. C. Oxidative Stress: An Essential Factor in the Pathogenesis of Gastrointestinal Mucosal Diseases. Physiol Rev. 2014;94:329-354. https://doi.org/10.1038/s41575-021-00440-6
CrossRef - Fan Y., Pedersen O. Gut Microbiota in Human Metabolic Health and Disease. Nat Rev Microbiol. 2021;19:55-71. https://doi.org/10.1038/s41579-020-0433-9
CrossRef - Choghakhori R., Abbasnezhad A., Hasanvand A., Amani R. Inflammatory Cytokines and Oxidative Stress Biomarkers in Irritable Bowel Syndrome: Association with Digestive Symptoms and Quality of Life. Cytokine. 2017;93:34-43. https://doi.org/10.1016/j.cyto.2017.05.005
CrossRef - Salminen S., Collado M. C., Endo A., Hill C., LebeerA., Quigley E. M. M., Sanders M. E., Shamir R., Swann J. R., Szajewska H., Vinderola G. The International Scientific Association of Probiotics and Prebiotics (ISAPP) Consensus Statement on the Definition and Scope of Postbiotics. Nat Rev Gastroenterol Hepatol. 2021. https://doi.org/10.1038/s41575-021-00440-6
CrossRef - Aguilar-Toalá J. E., Garcia-Varela R., Garcia H. S., Mata-Haro V., González-Córdova A. F., Vallejo-Cordoba B., Hernández- Mendoza A. Postbiotics: An Evolving Term Within the Functional Foods Field. Trends Food Sci Technol. 2018;75:105-114. https://doi.org/10.1016/j.tifs.2018.03.009
CrossRef - Zhao Y., Jiang Q. Roles of the Polyphenol–Gut Microbiota Interaction in Alleviating Colitis and Preventing Colitis-Associated Colorectal Cancer. Adv Nutr. 2020;12:546-565. https://doi.org/10.1093/advances/nmaa104
CrossRef - Davani-Davari D., Negahdaripour M., Karimzadeh I., Seifan M., Mohkam M., Masoumi A. J., Berenjian A., Ghasemi Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods. 2019;8:92-119. https://doi.org/10.3390/foods8030092
CrossRef - Contesini F. J., de Lima E. A., Mandelli F., Borin G. P., Alves R. F., Terrasan C. R. F. Carbohydrate Active Enzymes Applied in the Production of Functional Oligosaccharides, In: Encyclopedia of Food Chemistry. Oxford: Academic Press; 2019:30-34. https://doi.org/10.1016/B978-0-08-100596-5.21639-9
CrossRef - Dong Y., Xu M., Chen L., Bhochhibhoya A. Probiotic Foods and Supplements Interventions for Metabolic Syndromes: A Systematic Review and Meta-Analysis of Recent Clinical Trials. Ann Nutr Metabol. 2019;74:224-241. https://doi.org/10.1159/000499028
CrossRef - Dam B., Misra A., Banerjee S. Role of Gut Microbiota In Combating Oxidative Stress. In: Chakraborti S., Chakraborti T., Chattopadhyay D., Shaha C. Oxidative Stress in Microbial Diseases. Singapore: Springer; 2019:43-82. https://doi.org/10.1007/978-981-13-8763-0_4
CrossRef - Plaza-Diaz J., Ruiz-Ojeda F. J., Gil-Campos M., Gil A. Mechanisms of Action of Probiotics. Adv Nutr. 2019;10:S49-S66. https://doi.org/10.1093/advances/nmy063
CrossRef - Kruis W., Frič P., Pokrotnieks J., Lukáš M., Fixa B., Kaščák M., Kamm M. A., Weismueller J., Beglinger C. stole M. Maintaining Remission of Ulcerative Colitis with the Probiotic Escherichia coli Nissle 1917 Is As Effective As With Standard Mesalazine. Gut. 2004;53:1617-1623. https://doi.org/10.1136/gut.2003.037747
CrossRef - Chambers E. S., Preston T., Frost G., Morrison D. J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr Nutr Rep. 2018;7:198-206. https://doi.org/10.1007/s13668-018-0248-8
CrossRef - Den Besten G., Van Eunen K., Groen A. K., Venema K., Reijngoud D-J., Bakker B. M. The role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J Lipid Res. 2013;54:2325-2340. https://doi.org/10.1194/jlr.R036012
CrossRef - Ma G., Chen Y. Polyphenol Supplementation Benefits Human Health via Gut Microbiota: A Systematic Review via Meta-Analysis. J Funct Foods. 2020;66:103829-103845. https://doi.org/10.1016/j.jff.2020.103829
CrossRef - Kaja S., Payne A. J., Naumchuk Y. ,Koulen P. Quantification of Lactate Dehydrogenase for Cell Viability Testing Using Cell Lines and Primary Cultured Astrocyte. Curr Proto Toxicol. 2017;72:2.26.1-2.26.10. https://doi.org/10.1002/cptx.21
CrossRef - Leung H. H., Yau Y. F., Leung K. S, Lee Y. Y., Oger C., Durand T., Galano J. M., Loke W. M., Lee J. C. Y. Garlic Supplementation Modified Enzymatic Omega-6 Polyunsaturated Fatty Acid Oxidation in Mild Hypercholesterolemia. Eur J Lipid Sci Technol. 2019;121:1900069-190085. https://doi.org/10.1002/ejlt.201900069
CrossRef - Quek A., Mohd Zaini H., Nur Kartinee K., Sulaiman F., Rukayadi Y., Ismail A., Abidin Z. Z., Awang, K. Oxygen Radical Antioxidant Capacity (ORAC) and Antibacterial Properties of Melicope Glabra Bark Extracts and Isolated Compounds. PloS One. 2021;16: e0251534. https://doi.org/10.1371/journal.pone.0251534
CrossRef - Xiao F., Xu T., Lu B., Liu R. Guidelines for antioxidant assays for food components. Food Front. 2020;1:60-69. https://doi.org/10.1002/fft2.10
CrossRef - Assefa A.W., Keum Y.S., Saini R.K. A comprehensive study of polyphenols contents and antioxidant potential of 39 widely used spices and food condiments. J Food Measure Charact. 2018;12:1548-1555. https://doi.org/10.1007/s11694-018-9770-z
CrossRef - Leo L., Loong C., Ho X. L., Rahman M.F., Suan M.Y.T., Loke W.M. Occurrence of azo food dyes and their effects on cellular inflammatory responses. Nutrition. 2018;46:36-40. https://doi.org/10.1016/j.nut.2017.08.010
CrossRef - Loke W. M., Proudfoot J. M., Stewart S.McKinley A. J., Needs P. W., Kroon P. A., Hodgson J. M., Croft K. D. Metabolic Transformation has a Profound Effect on Anti-Inflammatory Activity of Flavonoids such as Quercetin: Lack of Association Between Antioxidant and Lipoxygenase Inhibitory Activity. Biochem Pharmacol. 2008;75:1045-1053. https://doi.org/10.1016/j.bcp.2007.11.002
CrossRef - Chang H. M., Foo H. L. , Toh T. C., Lim E. T. C., Abdul Mutalib N. E. Comparative Studies of Inhibitory and Antioxidant Activities, and Organic Acids Compositions of Postbiotics Produced by Probiotic Lactiplantibacillus plantarum Strains Isolated from Malaysian Foods. Front Vet Sci. 2020;7:1-14. https://doi.org/10.3389/fvets.2020.602280
CrossRef - Tian T. , Wang Z. , Zhang J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxid Med Cellul Longev. 2017. https://doi.org/10.1155/2017/4535194
CrossRef - Huang D., Ou B., Prior R. L. The Chemistry Behind Antioxidant Capacity Assays. J Agric Food Chem. 2005;53:1841-1856. https://doi.org/10.1021/jf030723c
CrossRef - Zheng Y. Z. , Deng G., Liang Q., Chen D-F., Guo R., Lai R-C. Antioxidant Activity of Quercetin And Its Glucosides From Propolis: A Theoretical Study. Sci Rep. 2017;7:7543-7554. https://doi.org/10.1038/s41598-017-08024-8
CrossRef - Robaszkiewicz A. A., Balcerczyk A., Bartos G. Antioxidative and Prooxidative Effects of Quercetin On A549 Cell. Cell Biol Int. 2007;31:1245-1250.
CrossRef - Yarahmadi A., Zal F., Bolouki A. Protective Effects Of Quercetin On Nicotine Induced Oxidative Stress In ‘HepG2 Cells’. Toxicol Mech Methods. 2017;27:609-614. https://doi.org/10.1080/15376516.2017.1344338
CrossRef - Sun L., Xu G., Dong Y., Li M., Yang L., Lu W.. Quercetin Protects Against Lipopolysaccharide-Induced Intestinal Oxidative Stress in Broiler Chickens Through Activation Of NRF2 Pathway. Molecules. 2020;25:1053-1065. https://doi.org/10.3390/molecules25051053
CrossRef - Amorati R., Baschieri A., Cowden A., Valgimigli L. The Antioxidant Activity Of Quercetin In Water Solution. Biomimetics. 2017;2:9-22. https://doi.org/10.3390/biomimetics2030009
CrossRef - Kumar Singh A., Cabral C., Kumar R.,Ganguly R., Kumar Rana H., Gupta A., Rosaria Lauro M., Carbone C., Reis F., Pandey A. K. Beneficial Effects Of Dietary Polyphenols On Gut Microbiota And Strategies To Improve Delivery Efficiency. Nutrients. 2019;11:2216-2237. https://doi.org/10.3390/nu11092216
CrossRef - Huang W., Guo H., Deng X, Zhu T. T., Xiong J-F., Xu Y-H., Xu, Y. Short-Chain Fatty Acids Inhibit Oxidative Stress and Inflammation in Mesangial Cells Induced by High Glucose and Lipopolysaccharide. Exp Clin Endocrinol Diabetes. 2017;125:98-105. https://doi.org/10.1055/s-0042-121493
CrossRef - Yuan X., Wang L., Bhat O.nM., Lohner H., Li P-L. Differential Effects of Short Chain Fatty Acids on Endothelial NLRP3 Inflammasome Activation and Neointima Formation: Antioxidant Action of Butyrate. Redox Biol. 2018;16:21-31. https://doi.org/10.1016/j.redox.2018.02.007
CrossRef - Almeida A. F., Borge G. I. A., Piskula M., Tudose A., Tudoreanu L., Valentová K., Williamson G., Santos C. N. Bioavailability Of Quercetin In Humans With A Focus On Interindividual Variation. Comp Rev Food Sci Food Safety. 2018;17:714-731. https://doi.org/10.1111/1541-4337.12342
CrossRef - Fuentes J., de Camargo A. C., Atala E.,Gotteland M., Olea-Azar C., Speisky H. Quercetin Oxidation Metabolite Present In Onion Peel Protects CACO-2 Cells Against The Oxidative Stress, NF-ĸB Activation, And Loss of Epithelial Barrier Function Induced By NSAIDs. J Agric Food Chem. 2021;69:2157-2167. https://doi.org/10.1021/acs.jafc.0c07085
CrossRef - Fernández J., Redondo-Blanco S., Gutierrez-del-Rio I., Miguelez E. M., Villar C. J., Lombo F. Colon Microbiota Fermentation of Dietary Prebiotics Towards Short-Chain Fatty Acids And Their Roles As Anti-Inflammatory And Antitumour Agents: A Review. J Funct Foods. 2016;25:511-522. https://doi.org/10.1016/j.jff.2016.06.032
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






