Introduction
Processed potato products are of the most popular foods and represent a large part of the food industry. At the same time improving the quality and safety of foods during processing is a never ending effort and an obligation to the consumers. Within this context of food quality and safety in recent years the acrylamide has been added to the list of harmful food substances. The formation of acrylamide during cooking at high temperature of unprocessed plant products was reported in 20021,2 and the presence of acrylamide in food is now recognized as a difficult problem for the food industry. The polymeric acrylamide substance has been characterized as neurotoxic and potentially carcinogenic and its formation is necessary to be prevented. Acrylamide is formed in foods mainly after heat treatment from the reaction of proteins with sugars as one of the many byproducts of the Mailard reaction in fried or grilled but not boiled foods.3,4 Many studies have already been done on a number of possible factors implicated in acrylamide formation in order to identify ways of prevention. As part of these studies, genetic background and cultivation methods have also been used whether they can affect the process of acrylamide formation.
The main food ingredients that lead to acrylamide formation in conditions of high heat (Mailard reaction) is free asparagine which reacts with reducing sugars.5 Many plants contain relatively high concentrations of free asparagine compared with other amino acids, indicating that plants use free asparagine for purposes such as the transport and storage of nitrogen.6 The genes encoding participating enzymes for asparagine and reducing sugars biosynthesis are possible targets for genetic modification to control acrylamide formation. The groups of genes SnRK1 and GCN2 are targets for genetic changes that affect the final formation of acrylamide by the control of reducing sugars and asparagine. Transgenic lines over expressing SnRK1 contain up to 83% less glucose and 30% more starch.7 Over expression of GCN2, has been shown to significantly reduce the expression of the asparagine synthetase gene. GCN2 gene expression is activated at low concentrations of free acid and the synthesized protein kinase acts by phosphorylating the alpha subunit of the eukaryotic translation initiation factor (eIF2), thereby reducing the rate of protein synthesis and free asparagine. The mechanism has been less studied compared with similar processes in fungi and further research is required.8
The above described approaches include genetic modification of plants banned in many countries as for example in Europe. For a better understanding of these two factors and their contribution to prevent acrylamide formation utilizing cultivation techniques, the current study was carried out. The aim of this study was the effect of two gene groups SnRK1 and GCN2 in two potato varieties Lady Rosetta and Spunta cultivated at different levels of nitrogen fertilization.
Materials and Methods
The effect of nitrogen fertilization in the expression of metabolic regulators SNRK1 and GCN2 genes, that control carbohydrate and protein synthesis and thus affecting the formation of acrylamide in two potato varieties Spunta and Lady Rosetta fried in corn oil was studied. Two levels of nitrogen were used during potato cultivation [treatment 1: N1=300mg/kg of soil, treatment 2: N2=600mg/kg of soil)] and were examined in relation to mRNA expression of the genes SnRK1 and GCN2 and acrylamide formation. Potato tubers were received from the different fertilization treatments for each variety at the same growth stage. Tubers were washed in water, cleaned with 70% ethanol and dried. For each sample case half of the potato tubers were homogenized and stored with the rest of the tubers at -70ºC. Homogenates were further analyzed for mRNA expression for the genes SnRK1 and GCN2, reducing sugars, total protein and while the rest was tested for acrylamide formation after frying.
Isolation of mRNA and Real Time PCR
For analysis of the mRNA expression of SnRK1 and GCN2, genetic material was isolated with the kit Total RNA isolation from plant (NucleoSpin® RNA Plant, Macherey-Nagel GmbH & Co. KG, Germany). In the isolated mRNA in order to compare expression levels of the SnRK1 and GCN2 genes, relative quantification of the mRNA fractions of the two genes was performed using the Kappa SybrTM Fast One step qRT-PCR kit (Kappa Biosystems, Boston, USA). The produced cDNA was processed in two step Real Time PCR with denaturation step at 95 oC for 3 sec and anneal/extension step at 60 oC for 30 sec for 35 cycles. Primer design was based on the analysis of sequences retrieved from the EMBL-EBI nucleic acid database. Results for all primers per case were analyzed in an Excel sheet and Ct values were averaged and compared. Ct values were indicative of the expression levels for the certain genes among different treatments.
Total protein, reducing sugars and acrylamide determination
Total soluble protein content was quantified with Bradford reagent (Sigma) according to the manufacturer’s protocol. The absorption of the dye was measured at 595nm. Quantification was expressed as g/dry weight and was based on a standard curve from a series dilution of BSA protein (Sigma).
Reducing sugars were quantified with Dinitrosalicylic acid (DNS) which reacts with reducing sugars to form 3-amino-5-nitrosalicylic acid .9 The absorbance of the samples was measured at 535 nm in a U-1500 spectrophotometer (HITACHI, Japan) using a glucose standard curve.
Acrylamide was determined with the Morinaga Acrylamide EIA kit according to manufacturer’s protocol. It enables simultaneous determinations of the acrylamide content in multiple test food samples within a short period of time (approx.4 h except for sample extraction) based on ELISA measurement.
Results
Results from this study are presented and analyzed in order to understand and control acrylamide formation with varying fertilization levels. Analysis involved RNA expression levels of genes SnRK1 and GCN2. In Figure 1 a representative sample from Real time PCR products analyzed on a 2% agaroze gel for the two potato varieties Spunta and Lady Rosetta is provided using 3 different pairs of primers per gene of interest. Analysis of gene expression was based on relative comparison of Ct values obtained from the Real Time PCR.
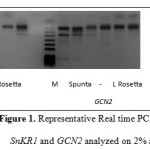 |
Figure 1: Representative Real time PCR products for SnKR1 and GCN2 analyzed on 2% agaroze gel. |
Figures 2 and 3 show the relative mRNA expression levels of SnRK1 and GCN2 genes for the two varieties Spunta and Lady Rosetta under different fertilization treatments, respectively. It is observed that for both varieties reduction of nitrogen fertilization (level 1) resulted in an increase in the expression of both genes in comparison to their expression with higher amounts of nitrogen fertilization (level 2).
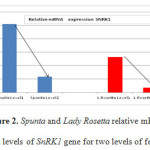 |
Figure 2: Spunta and Lady Rosetta relative mRNA expression levels of SnRK1 gene for two levels of fertilization. |
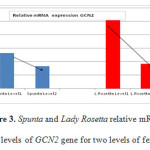 |
Figure 3: Spunta and Lady Rosetta relative mRNA expression levels of GCN2 gene for two levels of fertilization. |
Figures 4 and 5 show the relative mRNA expression levels of SnRK1 gene in relation to the amount of reducing sugars and acrylamide formation after frying for the two varieties Spunta and Lady Rosetta under different fertilization treatments. It is observed that for both verities a reduction in nitrogen fertilization (level 1) when compared with higher amounts of nitrogen fertilization (level 2), caused an increase in relative mRNA expression levels of SnRK1 gene, an increase in reducing sugars, but a reduction in acrylamide formation.
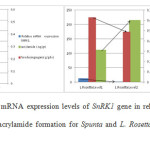 |
Figure 4, 5: Relative mRNA expression levels of SnRK1 gene in relation to the amount of reducing sugars and acrylamide formation for Spunta and L. Rosetta variety after frying in corn oil. |
Figures 6 and 7 show the relative mRNA expression levels of GCN2 gene in relation to protein content and acrylamide formation after frying for the two varieties Spunta and Lady Rosetta under different fertilization treatments. It is observed that for both varieties a reduction in nitrogen fertilization (level 1) when compared with higher amounts of nitrogen fertilization (level 2), caused an increase in relative mRNA expression levels of gene GCN2, a reduction in reducing sugars and a reduction in acrylamide formation.10
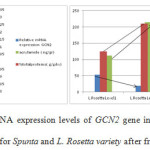 |
Figure 6, 7: Relative mRNA expression levels of GCN2 gene in relation to protein content and acrylamide formation for Spunta and L. Rosetta variety after frying in corn oil. |
Discussion
Summarizing the presented data it is highlighted that reduced nitrogen fertilization in all cases had a positive effect in acrylamide formation as it was lower when compared to acrylamide produced at higher nitrogen levels (twice the nitrogen level 1). Both genes seem to have a positive effect at low nitrogen levels in acrylamide formation. While low nitrogen caused an increase in expression of SnRK1, reducing sugars did not decrease indicating that over expression could be considered necessary for reducing sugars decrease as it happens when cloning and over expressing SnRK1 or in nutrient stress conditions. The reduced acrylamide levels despite increased reducing sugars at low nitrogen levels are attributed mainly to the reduction of free amino acids, specifically asparagine, in relation to GCN2 gene expression which is activated at low nitrogen levels and causes reduction of free asparagine.
Acknowledgments
This work was funded by the European Social fund and National Resources through Research Program Archimedes III.
References
- Tareke, E., P. Rydberg, P. Karlsson, S. Eriksson, and M. Tornqvist, ‘Analysis of acrylamide, a carcinogen formed in heated foodstuffs’, J. Agric. Food Chem. vol. 50, 2002, pp. 4998–5006.
CrossRef - Halford, N.G., T.Y. Curtis, N. Muttucumaru, J. Postles, J.S. Elmore, D.S. Mottram, ‘The acrylamide problem: a plant and agronomic science issue’, J. Experiment Bot. vol. 63 (8), 2012, pp. 2841-2851.
CrossRef - Mottram, D.S., B.L. Wedzicha, and A.T. Dodson, ‘Acrylamide is formed in the Maillard reaction’ Nature, vol. 419, 2002, pp. 448 – 449.
CrossRef - Stadler, R.H., I. Blank, N. Varga, F. Robert, J. Hau, P.A Guy, M.C. Robert, and S. Riediker, ‘Acrylamide from Maillard reaction products’ Nature, vol. 419, 2002, pp. 449–450.
CrossRef - Curtis, T. Y., N. Muttucumaru, P. R. Shewry, M. A. Parry, S. J. Powers, J. S. Elmore, D. S. Mottram, S. Hook, and N. G. Halford, ‘Evidence for genetic and environmental effects on free amino acid levels in wheat grain: implications for acrylamide formation during processing’, J. Agric. Food Chem., vol. 57, 2009, pp. 1013–1021.
CrossRef - Lea, P. J., L. Sodek, M. A. Parry, P. R. Shewry, and N. G. Halford, ‘Asparagine in plants’ Ann. Appl. Biol., vol. 150, 2007, pp. 1– 26.
CrossRef - McKibbin, R.S., N. Muttucumaru, M.J. Paul, S.J. Powers, M.M. Burrell, S. Coates, P.C. Purcell, A. Tiessen, P. Geigenberger, and N.G. Halford, ‘Production of high-starch, low-glucose potatoes through over expression of the metabolic regulator SnRK1’. Plant Biotechnol. J. vol. 4, 2006, pp. 409–418.
CrossRef - Halford, N., and I. Prosser, ‘Over-expression of GCN2-type protein kinase in plants’ Patent WO 2013038202 A2, 2013.
- Miller, D.R. ‘Use of dinitrosalicylic acid reagent for determination of reducing sugar’. Anal. Chem. Vol. 31, 1959, pp. 426–428.
CrossRef - Argyropoulos, D., Ch. Psallida and T. Varzakas. 2015. The effect of Nitrogen fertilisation and metabolic regulators SNRK1, GCN2 on the formation of acrylamide in two potato varieties (Spunta and Lady Rosetta) fried in corn oil. Poster presentation in session on manipulating postharvest physiology to improve food quality. IJFST 50th celebration conference: the future of food innovation, nutrition and technology. 17-19 February 2015, Lincoln University, New Zealand, P19.

This work is licensed under a Creative Commons Attribution 4.0 International License.








