
Introduction
Spirulina, a filamentous cyanobacterium belonging to the genus Arthrospira,1 exhibits remarkable versatility as a microorganism, boasting a rich history of human utilization.2 Flourishing in diverse environments, including alkaline freshwater and saltwater, Spirulinahas been utilizing as a dietary staple in various cultures worldwide.3 Recent scientific investigations have cast a spotlight on Spirulina is potential as a source of folic acid.4 This microorganism possess inherent folate production capabilities according to study done by Wang and others.5 Furthermore, when subjected to co-cultivation with folate-producing bacteria, Spirulina has demonstrated the capacity to enhance folic acid production, resulting in a synergistic effect on folate synthesis.6 These discoveries unveil promising pathways for the creation of economical and environmentally sustainable methodologies for folic acid synthesis.
Folate, often known as vitamin B9, is an essential nutrient that plays a variety of roles in important biological processes in the human body. Its significance extends from its pivotal involvement in DNA synthesis and cell division to its crucial contribution to fetal neural tube development.7,8 However, despite its fundamental importance, the global challenge of folate deficiency persists, affecting specific populations and necessitating the exploration of innovative strategies for its amelioration.
Previous research has gone extensively into the multiple repercussions of folate deficiency and the many techniques used to address this worldwide health problem. Notably, low folate levels have been linked to an increased risk of various health problems. One of the most noticeable outcomes of folate deficiency is the incidence of neural tube abnormalities (NTDs) in growing fetuses.9 The insufficiency of maternal folate intake during pregnancy has been linked to an increased risk of NTDs, severe malformations of the brain and spinal cord.10 Additionally, diminished folate levels have been implicated in the development of various conditions, encompassing cardiovascular diseases, specific cancer types (such as colorectal, pancreatic, and cervical cancer), and neurodegenerative disorders like Alzheimers disease.11 These investigations collectively underscore the critical nature of folate, especially during periods characterized by rapid growth, thereby emphasizing its indispensable significance in fetal development and overall health.
In response to the persistent challenge posed by folate deficiency, we embarked on a research endeavor aimed at investigating an innovative approach to address this issue. Researchers are exploring the possibility of microbial fermentation to produce natural folate. Several biotechnological and chemical methods are used to make folic acid in high production scale, Bacillus subtilis has been modified to boost the synthesis of folate by co-culturing strategy. Microbial manufacturing is a sustainable and cost-effective technique to manufacture an appropriate blend of folate vitamins. Specifically, our study focused on examining the synergistic interaction between Spirulina, a microalga known for its rich nutritional content, and two folic acid-producing bacteria strains, Bacillus subtilis-1 and Bacillus subtilis-2, with the primary objective of augmenting folate production. The rationale behind this investigative pursuit stemmed from the remarkable nutritional profile exhibited by Spirulina, coupled with the inherent folate production capabilities exhibited by the chosen bacterial strains. Our methodological approach entailed the separate cultivation of Spirulina and the two strains of Bacillus subtilis, allowing for the optimization of growth conditions tailored to each organisms’ unique requirements. Subsequently, co-culture experiments were conducted, bringing together Spirulina with Bacillus subtilis-1 and Bacillus subtilis-2, respectively, to explore their combined potential for folate production. The outcomes showed the profound impact of microbial cooperation on nutrient production. The implications of this research venture extend beyond the laboratory, offering insights into the collaborative potential of microorganisms and holding significant promise for addressing global challenges related to folate deficiencies. The synergistic association between Spirulina and folic acid-producing Bacillus subtilis strains presents a promising approach for the development of functional foods enriched with folate, thus contributing to improved health outcomes. Moreover, the study’s objective was to harness a microalgae-heterotrophic bacteria co-culture, specifically between Spirulina and Bacillus subtilis, to boost folate production, aiming for functional foods with enhanced nutritional value.Ultimately, these findings offer innovative, sustainable, and eco-friendly approaches to meet the ever-growing nutritional demands of our global populace.
Materials and Methods
The cyanobacteria and bacteria that showed the highest folate production capabilities were selected for co-culture experiments. The co-culture experiments were conducted using a two-step process. In the first step, the folic acid producing bacteria and Cyanobacteria Specifically Spirulina were cultured separately in a nutrient-rich medium. In the second step, the folic acid producing cyanobacteria were added to the bacterial culture at different time intervals (24 Hours). The co-culture experiments were conducted under different environmental conditions, and the folate production was measured at different time points using standard microbiological techniques. The folate production was quantified using Phosphate Buffer Extraction Method.
Study Area
Bacterial strains from yogurt and Spirulinasp. from Usterzai, Kohat, KPK, Pakistan, were previously identified in a lab project. The experiment was conducted at the Laboratory of Molecular Ecology and Conservation, Department of Biotechnology and Genetic Engineering, Kohat University of Science and Technology, Kohat, KP, Pakistan.
Culture Cultivation for Consortia
In this study, the cultivation techniques12 were followed to establish large-scale and sustainable cultivation of microalgae. These techniques are crucial for ensuring optimal growth and productivity of microalgae cultures. They involve providing suitable growth conditions such as temperature, light intensity, humidity and pH in a growth chamber specially design for this experiment. The cultivation setup consisted of culture flasks, depending on the scale of the experiment. Spirulina sp. is cultured in BG-11 media, while bacterial strains for the study are maintained in MRS broth media.
Furthermore, the methods13 were adopted to establish microalgal-bacterial interactions in aquaculture systems. The microalgae-bacteria consortia were carefully prepared by inoculating the microalgae culture with the selected bacteria under controlled laboratory conditions like pH -10, temperature-22°C and light intensity 2400lux. This approach aimed to promote a synergistic relationship between microalgae and bacteria, resulting in increased folate production.
Specific Growth Rate Measurement of Folate-Producing Bacteria and Spirulina for Enhanced Folate Production
To determine the specific growth rate of folic acid-producing bacteria and Spirulina sp., a widely employed spectrophotometric method was utilized for folic acid-producing bacteria14 and for Spirulina sp.15
For folic acid-producing bacteria, the samples were analyzed at regular intervals (24 Hours), and their optical density (OD) was measured at a specific wavelength of 580nm using a spectrophotometer.14 The OD measurement at 580nm is commonly used to estimate the cell concentration in bacterial cultures, as it correlates with the microbial biomass present in the sample. The increase in OD over time allowed us to determine the specific growth rate of the folic acid-producing bacteria accurately.
For Spirulina sp., the spectrophotometric method was adopted for OD measurements at 580nm.15 This technique facilitated the calculation of Spirulina’s specific growth rate by analyzing OD value changes.
Dry Weight Analysis of Folate-Producing Bacteria and Spirulina for Enhanced Folate Production
For dry weight analysis, folate-producing bacteria and Spirulina were co-cultured to their respective growth phases. Daily, from day 1 to day 12, 1.5 ml samples were centrifuged to separate biomass from the medium. The supernatant was discarded, and the pellet washed to remove media residues. The dry weight of this cleaned pellet was measured on a balance to assess biomass productivity and growth performance, providing essential data for calculating specific growth rates and enhancing folate production.16
Harmonious Biosynthesis: Exploring Folate Quantification and Symbiotic Insights inMicroalgal-Bacterial Cultures
Folate levels in microalgal-bacterial cultures were determined by spectrophotometric analysis.17 This entailed extracting folate from microalgal biomass via the phosphate buffer method and measuring absorbance at 282nm with a C-7200S double UV-visible spectrophotometer. A standard folate curve aided in precise measurement, with absorbance readings compared to the curve for calculating folate concentration. The symbiotic interaction between algae and bacteria in vitamin B12 acquisition, which is relevant to folate generation, was investigated.18 This study investigated the process of folate synthesis in a microalgal-bacterial culture, with the goal of increasing folate production through symbiosis.
Statistical Analysis
The current data were presented in the form of average and SD. To know the difference between the groups the t tests, tests were applied by using Statistics version 9. The P values were considered significantly const.
Results
Synergistic Growth and Folate Production of Bacillus subtilis-1
During the log phase (Days 2-5), Bacillus subtilis-1 exhibited a specific growth rate of 0.269±0.321 µ, biomass productivity of 0.001±0.000, and folate production of 0.043±0.003 µg/mL, suggesting active growth and metabolite synthesis. In the stationary phase (Days 6-11), a decline in specific growth rate to -0.036±0.036 µ was observed alongside decreased biomass productivity (-0.001±0.000) and slightly reduced folate production (0.034±0.005 µg/mL).Table 1, indicating a shift towards metabolic equilibrium. Fig. 1.
Table 1: Average Growth Dynamics and Folate Production in different phases of Bacillus subtilis-1
|
Phases |
Treatment (Days) | Specific Growth Rate (µ) | Biomass Productivity |
Folate production (µg/mL) |
|
Lag |
1 | 0.000±0.000 | 0.000±0.000 | 0.000±0.000 |
|
Log |
2-5 | 0.269±0.321 | 0.001±0.000 | 0.043±0.003 |
| Stationary | 6-11 | -0.036±0.036 | -0.001±0.000 |
0.034±0.005 |
| Decline | 12 | -0.069±0.000 | -0.001±0.000 |
0.026±0.000 |
 |
Figure 1: Dynamics for Bacillus substilis-1 |
Unveiling Growth Dynamics and Folate Production of Bacillus Substilis-2
For Bacillus Subtilis-2, the organism showed robust growth in the log phase (Days 2-3) with a specific growth rate of 0.446±0.399 µ, minimal biomass productivity (0.001±0.000), and initiating folate production at 0.041±0.002 µg/mL. Entering the stationary phase (Days 4-9), growth significantly slowed (0.010±0.067 µ), and biomass productivity slightly declined (-0.001±0.002), yet folate production remained stable at 0.041±0.005 µg/mL. The decline phase (Days 10-12) witnessed a decrease in growth rate (-0.072±0.029 µ) Table 2, further reduced biomass productivity (-0.002±0.002), and a slight reduction in folate production to 0.034±0.001 µg/ml Fig.2.
Table 2: Average Growth Dynamics and Folate Production in different phases of Bacillus subtilis-2
|
Phases |
Treatment (Days) | Specific Growth Rate (µ) | Biomass Productivity | Folate production (µg/mL) |
|
Lag |
1 | 0.000±0.000 | 0.000±0.000 | 0.000±0.000 |
| Log | 2-3 | 0.446±0.399 | 0.001±0.000 |
0.041±0.002 |
|
Stationary |
4-9 | 0.010±0.067 | -0.001±0.002 |
0.041±0.005 |
| Decline | 10-12 | -0.072±0.029 | -0.002±0.002 |
0.034±0.001 |
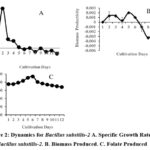 |
Figure 2: Dynamics for Bacillus substilis-2. |
Revealing Growth Dynamics and Folate Production of Spirulina
For Spirulina, during the lag phase (Days 1-4), a specific growth rate of 0.212±0.182 µ was observed alongside biomass productivity of 0.041±0.028 and negligible folate production (0.000±0.000 µg/mL). In the subsequent log phase (Days 5-7), growth continued albeit at a slower rate, with a specific growth rate of 0.126±0.051 µ, biomass productivity of 0.026±0.009, and a minimal increase in folate production (0.001±0.001 µg/mL). During the stationary phase (Days 6-11), growth further slowed (0.045±0.045 µ), but biomass productivity increased slightly to 0.036±0.008, and folate production rose to 0.002±0.000 µg/mL. Finally, in the decline phase (Day 12), growth ceased (0.000±0.035 µ) Table 3, biomass productivity remained relatively constant (0.032±0.000), and folate production reverted to negligible levels (0.000±0.000 µg/mL) Fig. 3.
Table 3: Average Growth Dynamics and Folate Production in different phases of Spirulina
|
Phases |
Treatment (Days) | Specific Growth Rate (µ) |
Biomass Productivity |
Folate production (µg/mL) |
|
Lag |
1-4 | 0.212±0.182 | 0.041±0.028 | 0.000±0.000 |
| Log | 5-7 | 0.126±0.051 | 0.026±0.009 |
0.001±0.001 |
|
Stationary |
6-11 | 0.045±0.045 | 0.036±0.008 | 0.002±0.000 |
| Decline | 12 | 0.000±0.035 | 0.032±0.000 |
0.000±0.000 |
 |
Figure 3: Dynamics for Spirulina |
Unveiling Synergistic Interplay between Spirulina and Bacillus Subtilis-1 for Enhanced Folate Production
In co-culture, Spirulina and Bacillus Substilis-1 showed no growth or biomass productivity during the lag phase (Day 1), but an unusual initial folate production of 0.468±0.000 µg/mL was observed. During the log phase (Days 2-7), they exhibited a specific growth rate of 0.077±0.076 µ, minimal biomass productivity (0.001±0.002), and an increased folate production of 0.519±0.082 µg/mL. However, in the stationary phase (Days 8-11), a decrease in specific growth rate to -0.080±0.024 µ was noted alongside a slight decline in biomass productivity (-0.001±0.001) and a marginal reduction in folate production to 0.502±0.033 µg/mL Table 4. The decline phase (Day 12) saw all parameters reverting to zero, indicating cessation of growth and metabolite production Fig. 4
Table 4: Average Growth Dynamics and Folate Production in different phases of Spirulina co-cultured with Bacillus substilis-1
|
Phases |
Treatment (Days) | Specific Growth Rate (µ) | Biomass Productivity |
Folate production (µg/mL) |
|
Lag |
1 | 0.000±0.000 | 0.000±0.000 | 0.468±0.000 |
| Log | 2-7 | 0.077±0.076 | 0.001±0.002 |
0.519±0.082 |
|
Stationary |
8-11 | -0.080±0.024 | -0.001±0.001 | 0.502±0.033 |
| Decline | 12 | 0.000±0.000 | 0.000±0.000 |
0.000±0.000 |
 |
Figure 4: Synergistic Dynamics for Spirulina co-cultured with Bacillus substilis-1. |
Unveiling the Synergistic Dynamics of Spirulina and Bacillus substilis-2 for Folate Enhancement
The co-culture of Spirulina with Bacillus subtilis-2 showed no initial growth or biomass productivity but had an early folate production of 0.468±0.000 µg/mL during the lag phase. The log phase witnessed an increase in specific growth rate to 0.188±0.147 µ and slight biomass productivity (0.001±0.002), with folate production rising to 0.519±0.074 µg/mL. In the stationary phase, a decline in specific growth rate (-0.029±0.043 µ) and a decrease in biomass productivity (-0.001±0.001) were observed, yet folate production remained relatively stable at 0.502±0.033 µg/mL Table 5. By the decline phase, all measured activities ceased, marking the end of the co-culture’s growth cycle Fig. 5.
Table 5: Average Growth Dynamics and Folate Production in different phases of Spirulina co-cultured with Bacillus substilis-2
|
Phases |
Treatment (Days) | Specific Growth Rate (µ) | Biomass Productivity | Folate production (µg/mL) |
| Lag | 1 | 0.000±0.000 | 0.000±0.000 |
0.468±0.000 |
|
Log |
2-7 | 0.188±0.147 | 0.001±0.002 | 0.519±0.074 |
| Stationary | 8-11 | -0.029±0.043 | -0.001±0.001 |
0.502±0.033 |
|
Decline |
12 | 0.000±0.000 | 0.000±0.000 |
0.000±0.000 |
 |
Figure 5: Synergistic Dynamics for Spirulina co-cultured with Bacillus substilis-2. |
Discussion
The results presented in this study reveal significant insights into the growth dynamics and folate production of Bacillus subtilis-1, Bacillus Substilis-2, and Spirulina. The distinct growth phases observed in each microorganism played crucial roles in shaping their folate synthesis capabilities. Understanding these growth dynamics and their impact on folate production is essential for harnessing their potential in various applications. The exponential phase observed in Bacillus subtilis-1 showcased rapid growth and heightened folate synthesis. This phase is characterized by robust metabolic activities and efficient resource utilization, contributing to increased biomass and folate production. Similar findings have been reported in other folate-producing bacteria, emphasizing the significance of the exponential phase in enhancing folate synthesis.19
During the stationary phase of Bacillus subtilis-1, a balance between growth and sustained folate production was observed. This phase demonstrated stable growth conditions and a continued rise in folic acid production, indicating a consistent folate synthesis under favorable growth conditions. Other studies on Bacillus species have reported similar findings, suggesting that stationary phase cultures maintain folate production rates despites lower growth.18, 20
The decline phase in Bacillus subtilis-1 revealed the importance of optimal conditions for preserving folate synthesis. As the culture faced unfavorable conditions and nutrient limitation, both specific growth rate and folate production declined significantly. This reduction in metabolic activities and folate synthesis during the decline phase is consistent with observations in another Bacillus cultures.21
Moving on to Bacillus Substilis-2, the exponential phase exhibited a surge in folate synthesis despite a decline in growth rate. This phenomenon suggests a beneficial interplay between Bacillus Substilis-2 and the folate biosynthetic pathways, leading to enhanced folate production. The simultaneous increase in folic acid production and stability in biomass during this phase imply efficient nutrient utilization and metabolic activities.22
In the stationary phase of Bacillus Substilis-2, a balance between growth and folate production was maintained. The specific growth rate slightly decreased, but folate production remained relatively stable. This phase represents a period of adaptation and stabilization in the co-culture, allowing for the optimization of resource utilization and maintaining a balance between growth and folate.23
The decline phase in Bacillus Substilis-2 demonstrated reduced growth, metabolic activities, and folate production. The decline in specific growth rate and folate production during this phase can be attributed to various factors, such as nutrient depletion, altered microenvironment, or potential competition within the co-culture.24
Spirulina exhibited unique growth dynamics during the experiment. The exponential phase of Spirulina is characterized by rapid growth and increased metabolic activity, which plays a significant role in enhancing folate synthesis.25 During the stationary phase of Spirulina, a balance between growth and folate production was observed. The specific growth rate decreased slightly, but the biomass remained constant, indicating a dynamic equilibrium between cell division and cell death. The steady rise in folate production during this phase suggests sustained folate synthesis under stable growth conditions.26
The co-cultivation of Spirulina with B. subtilis-1 and B. subtilis-2 revealed intriguing synergistic effects on folate production. Despite minimal growth and biomass productivity during the lag phase, an early and substantial increase in folate production was observed in both co-cultures. This suggests a potential cross-feeding or cooperative interaction between Spirulina and the Bacillus strains, enhancing folate biosynthesis. Furthermore, the fluctuations in specific growth rates and biomass productivity throughout the co-culture phases indicate dynamic metabolic interactions between the two organisms. Overall, the findings of this study shed light on the growth dynamics and folate production of the studied microorganisms, providing valuable insights into their potential applications in enhancing folate synthesis. Further research is warranted to explore the underlying mechanisms of the observed synergistic interactions and optimize the conditions for maximizing folate production in various industries.
Conclusion
Our research on Spirulina and folic acid-producing bacteria demonstrated synergistic interactions that increased folate synthesis across different growth stages. Co-culturing with Bacillus Subtilis-1 and Bacillus Subtilis-2, in particular, optimized growth and metabolic activities in the exponential phase while preserving cooperative contacts in the stationary phase for balanced symbiosis. The decline phase showed growth constraints, emphasizing the significance of understanding these dynamics for better folate synthesis. This study highlights the potential of microbial synergy for sustainable nutrient production, laying the framework for future co-culturing techniques.
Recommendation
The outcomes of this study call for further investigation and application of co-culturing systems to maximize folate production. As we face global concerns like as population growth and rising demand for key nutrients, using microorganisms’ collaborative capacity appears to be a possible road forward. The dance of growth and folate generation observed in Spirulina and its co-cultures demonstrates the beauty of microbial symbiosis. Moving forward, this understanding has the potential to inspire novel ideas and sustainable behaviors. Understanding and utilizing the synergistic interactions amongst microbes has the potential to revolutionize folate synthesis, contributing to a more nourished and thriving planet. The fascinating narrative of Spirulina and its microbial allies acts as a guiding ideal, stimulating study and collaboration for a healthier and more sustainable future.
Acknowledgement
Higher Education Commission of Pakistan and KUST, Kohat for providing facilities to conduct this research project.
Funding Sources
This work was supported by the Higher Education Commission of Pakistan through a project titled “Folate Production by Spirulina: Potential Nutritional and Therapeutic Supplements”. (Grant No. 9835).
Conflict of Interest
The authors do not have any conflict of interest.
Data Availability Statement
This statement does not apply to this article.
Ethics Statement
This research did not involve human participants, animal, subjects, or any material that requires ethical approval.
Informed Consent Statement
This study did not involve human participants, and therefore, informed consent was not required.
Permission to Reproduce Material from other Sources
Not applicable
Clinical Trial Registration
This research does not involve any clinical trials.
Author Contributions
- Maha Rehman: Conceptualization, Methodology, Analysis, Writing
- Rehan Naeem: Conceptualization, Methodology, Review & Editing
- Sunanda Biswas: Visualization, Writing – Review & Editing
- Maheen Sohail: Methodology, Analysis, Data Collection
- Sadeeq Khan: Writing – Review & Editing
- Muhammad Arif: Methodology, Analysis, Data Collection
- Nosheen Amjad: Writing – Review & Editing
- Saleha Tahir: Writing – Review & Editing
- Amina Mehmood: Writing – Review & Editing
References
- Sanchez M., Bernal-Castillo J., Rozo C., Rodríguez I. Spirulina(Arthrospira): an edible microorganism: a review. UniversitasScientiarum. 2003; 8(1):7-24.
- Rajvanshi M., Sagaram U. S., Subhash G. V., Kumar G. R. K., Kumar C., Govindachary S., Dasgupta, S. Biomolecules from microalgae for commercial applications. In Gayen K., Bhowmick T.K., Maity S.K. (Eds). Sustainable downstream processing of microalgae for industrial application. CRC Press. 2019;3-38.
CrossRef - Raja R., Coelho A., Hemaiswarya S., Kumar P., Carvalho I S., Alagarsamy A. Applications of microalgal paste and powder as food and feed: An update using text mining tool. Beni-SuefUniversity Journal of Basic and Applied Sciences. 2018; 7(4); 40-747.
CrossRef - Ge, Y., Kang, Y. K., Dong, L., Liu, L. H., & An, G. Y. (2019). The efficacy of dietary Spirulinaas an adjunct to chemotherapy to improve immune function and reduce myelosuppression in patients with malignant tumors. Translational cancer research, 8(4), 1065.
CrossRef - Wang X., Li Z., Su J., Tian Y., Ning X., Hong H., Zheng T. Lysis of a red-tide causing alga, Alexandrium tamarense, caused by bacteria from its phycosphere. Biological Control. 2010; 52 :123-130.
CrossRef - Woortman D V., Fuchs T., Striegel L., Fuchs., Weber N., Brück T B., Rychlik M. Microalgae a Superior Source of Folates: Quantification of Folates in Halophile Microalgae by Stable Isotope Dilution Assay. Front BioengBiotechnol. 2020;21(7):481.
CrossRef - Aslam F., Muhammad S. M., Aslam S., Irfan, J. A. Vitamins: key role players in boosting up immune response-a mini review. Vitamins & Minerals. 2017; 6(01).
- Kurowska K, Kobylińska M, Antosik K. Folic acid – importance for human health and its role in COVID-19 therapy. RoczPanstwZaklHig. 2023;74(2):131-141.
CrossRef - Wang X., Yu J., Wang, J. Neural Tube Defects and Folate Deficiency: Is DNA Repair Defective?. International Journal of Molecular Sciences. 2023; 24(3):2220.
CrossRef - Hasler M, Fideli ÜS, Susi A, Hisle-Gorman E. Examining the relationship between autism spectrum disorder and neural tube defects. CongenitAnom (Kyoto). 2023;63(4):100-108. doi: 10.1111/cga.12516.
CrossRef - Dwivedi M, Ghosh D, Saha A, Hasan S, Jindal D, Yadav H, Yadava A, Dwivedi M. Biochemistry of exosomes and their theranostic potential in human diseases. Life Sci. 2023;315:121369.
CrossRef - Wolkers H, Barbosa M., Kleinegris D. M. M., Bosma R., Wijffels R. H. In Microalgae:the green gold of the future? Large-scale sustainable cultivation of microalgae for the production of bulk commodities.Wageningen UR Food &Biobased Research, Wageningen. 2011.
- Natrah F. M. I., Bossier P., Sorgeloos P., Yusoff F. M., Defoirdt T. Significance of microalgal–bacterial interactions for aquaculture. Reviews in Aquaculture. 2013; 6(1).
CrossRef - Alves D. R., Maia de Morais S., Tomiotto-Pellissier F., Miranda-Sapla M. M., Vasconcelos F. R., Silva I. N. G. D.,Araujo de Sousa H., Assolini J. P., Conchon-Costa I., Pavanelli W R., Freire, F. D. C. O. Flavonoid composition and biological activities of ethanol extracts of CaryocarcoriaceumWittm., a native plant from Caatinga biome. Evidence-Based Complementary and Alternative Medicine. 2017. Article ID 6834218
CrossRef - Nayak P. K., Yang L., Brehm W., Adelhelm, P. From lithium‐ion to sodium‐ion batteries: advantages, challenges, and surprises. AngewandteChemie International Edition.2018;57(1):102-120.
CrossRef - Wicaksono, H. A., Satyantini, W. H., &Masithah, E. D. (2019, February). The spectrum of light and nutrients required to increase the production of phycocyanin Spirulinaplatensis. In IOP Conference Series: Earth and Environmental Science(Vol. 236, No. 1, p. 012008). IOP Publishing.
CrossRef - Tandon P., Jin Q., Huang L. A promising approach to enhance microalgaeproductivitybyexogenoussupplyof vitamins. Microbial Cell Factories. 2017;16:219.
CrossRef - Croft M. T., Lawrence A. D., Raux-Deery E., Warren M. J., Smith A. G. Algae acquire vitamin B12 through a symbiotic relationship with bacteria. Nature. 2005;438(7064):90-3.
CrossRef - Saubade F., Hemery Y. M., Guyot J. P., Humblot C. Lactic acid fermentation as a tool for increasing the folate content of foods. Critical Reviews in Food Science and Nutrition. 2017;57(18):3894-3910.
CrossRef - Scott J., Rébeillé F., Fletcher J. Folic acid and folates: the feasibility for nutritional enhancement in plant foods. Journal of the Science of Food and Agriculture.2000; 80(7):795-824.
CrossRef - Aragaw, W. W., Lee, B. M., Yang, X., Zimmerman, M. D., Gengenbacher, M., Dartois, V., Chui W.K., Jackson C.J., Dick, T. Potency boost of a Mycobacterium tuberculosis dihydrofolate reductase inhibitor by multienzyme F420H2-dependent reduction. Proceedings of the National Academy of Sciences.2021; 118(25): e2025172118.
CrossRef - Wu Z., Qiu S., Abbew A. W., Chen Z., Liu Y., Zuo J., Ge S. Evaluation of nitrogen source, concentration and feeding mode for co-production of fucoxanthin and fatty acids in Phaeodactylumtricornutum. Algal Research. 2022; 63 :102655.
CrossRef - Konstantinidis, D., Pereira, F., Geissen, E. M., Grkovska, K., Kafkia, E., Jouhten, P., Kim Y, Devendran S, Zimmermann M, Patil, K. R. Adaptive laboratory evolution of microbial co‐cultures for improved metabolite secretion. Molecular systems biology. 2021; 17(8):e10189.
CrossRef - Tong C. Y., Honda K., Derek C. J. C. A review on microalgal-bacterial Co-culture: The multifaceted role of beneficial bacteria towards enhancement of microalgal metabolite production. Environmental research. 2023:115872.
CrossRef - Garofalo C., Norici A., Mollo L., Osimani A., Aquilanti L. Fermentation of microalgal biomass for innovative food production. Microorganisms. 2022; 10(10):2069.
CrossRef - Slavov N., Budnik B. A., Schwab D., Airoldi E. M., vanOudenaarden, A. Constant growth rate can be supported by decreasing energy flux and increasing aerobic glycolysis. Cell reports. 2014; 7(3):705-714.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.






