
Introduction
Cardiovascular disease (CVD) affects the heart and blood vessels, and clinically manifests as myocardial infarction, heart failure, and stroke. It is the leading cause of death worldwide, with approximately 80% of deaths occurring in countries with low and middle incomes 1. The underlying cause of the disease is atherosclerosis, which develops over the years from fatty streaks to plaque formation in the presence of high levels of cholesterol 2,3. Cholesterol circulating throughout the body is supplied by both dietary cholesterol and endogenous cholesterol from liver synthesis, with the latter contributing to at least two-thirds of the total supply 4.
The key enzyme in cholesterol biosynthesis is 3-hydroxy-3-methylglutaryl coenzyme A (HMG-CoA) reductase, which is the target of a group of compounds called statins. Statins inhibit HMG-CoA reductase through competitive inhibition and effectively lower the blood cholesterol levels. As such, they are widely used in the treatment of hypercholesterolemia 5-7. However, although extensive research has been conducted on combination drug therapies to manage CVD, little work has been done on natural cholesterol-lowering compounds detected in some functional foods and nutraceuticals derived from plants. These foods are grouped into several types according to the mechanism of cholesterol reduction, with the HMG-CoA reductase inhibitor class identified as the most efficient in lowering blood cholesterol 8. Some bioactive compounds that inhibit HMG-CoA reductase include biflavonoids from tangerine peel extracts 9, diterpenes from the indigenous Indian plant Polyalthia longifolia 10, and phenolics from grapefruit peels 11.
In recent years, protein-derived bioactive peptides have been investigated as potential agents for reducing the risk factors for CVD 12-14. Bioactive peptides are short chains of amino acids encrypted within large protein molecules that exert beneficial health effects only when released from the intact protein through digestion or hydrolysis 12. Plant sources of such peptides are of interest as they are deemed more sustainable, cheaper, and environmentally-friendly alternatives than animal sources 15. Soybean peptides are one of the extensively studied plant peptides that have been reported to have antihypertensive and hypocholesterolemic activity and carry a US FDA-approved health claim for reduction of the risk of coronary heart disease (CHD), one of the CVDs, with regular consumption of the recommended quantity 16.
Lemongrass (Cymbopogon citratus Stapf.) as a traditional herb, has long been used in cooking, and is often consumed as an herbal tea. It is used primarily for the treatment of digestive and kidney problems, as well as other ailments, including hypertension, in the Philippines. Al Disi et al. (2016) 17 reviewed lemongrass as an anti-hypertensive herb, which was notably demonstrated through its vasorelaxation effects on constricted rat aortic rings. Lemongrass herbal tea has been reported to contain flavonoids, phenolic compounds, and citral, which are responsible for its observed health benefits 18-20 such as antioxidant 21,22, anti-inflammatory 23, antihypertensive 17,24,25, antidiabetic 26, and anti-cancer activities 27,28. In a study involving normotensive human subjects 29, lemongrass infusions administered to 105 test subjects lowered the blood pressure indices. However, although the authors implicated protein-derived peptides from lemongrass as possible antihypertensive agents, no experimental evidence has been presented to support this. Thus, this study determined the potential cholesterol-lowering effects of protein hydrolysates obtained from the digestion of lemongrass proteins, both in vitro and in an animal model. The results of this study will pave the way for a better understanding of the potential of protein hydrolysates and bioactive peptides from lemongrass as functional foods to prevent the risk of CVD.
Materials and Methods
Lemongrass plant materials
Lemongrass plants were collected on July 7-15, 2019 from Brgy. Baong, Alimodian, Iloilo, Philippines, and voucher specimen was deposited at the UST Herbarium (Certificate Acc. No. USTH 014150). The plants were washed, air-dried, then oven-dried at 50 – 60 °C until moisture content reached 5 – 6 %. The samples, consisting of leaf blade and leaf sheath, were ground into a fine powder and defatted with hexane at a ratio of 1:6 for 4 h before protein extraction.
Preparation of tannin-free protein extract (TFPE)
Proteins were extracted following the procedure described by Marques et al. (2015) 30 and Wang et al. (2017) 31 with modifications. Defatted lemongrass powder was dispersed in ultrapure water (18.2 MΩ.cm at 25 ⁰C) at a 1:10 w/v ratio, sonicated for 5 min at 20 ⁰C, and adjusted to pH 8.5 using 1 M NaOH. This suspension was agitated for 2.5 h, then centrifuged (Hettich Universal 320/320R) for 15 min at 8,000 rpm and 10 ºC, and the total protein in the supernatant was precipitated by adjusting the pH to 4 using 1 M HCl and collected by centrifugation for 15 min at 8,000 rpm and 10 ⁰C. The precipitate was further treated with polyamide (Fluka) to remove residual tannins, which would interfere with in vitro enzyme assays and in vivo gastrointestinal digestion. The resulting TFPE was then freeze-dried.
Digestion of proteins
Digestive juices were obtained from six male Sprague-Dawley rats weighing 200 – 250 g as described by Pivetta et al. (1981) 32 and Hosseinzadeh et al. (2002) 33, with modifications. Ethical considerations and housing conditions are described in the animal methods section (Section 2.7). After acclimatization, the animals were fasted overnight, anesthetized with Zoletil (Tiletamine-Zolazepam, 0.1 mg/kg body weight, BW), and immobilized by clamping the limbs. The abdominal area was dissected to locate the common bile duct. A polyethylene cannula was inserted into the duodenal lumen, and pancreatic juice was collected for one hour by spontaneous drainage into falcon tubes. The pylorus was then ligated, the stomach contents were quickly drained into tubes, and the animals were euthanized by Zoletil overdose. The pancreatic juice was freeze-dried, and the gastric juice was further centrifuged to remove the sediments and freeze-dried.
TFPE was digested in vitro by simulating GI conditions as described by Capriotti et al. (2015) 34. Briefly, 1 mg of TFPE was dissolved in 1 mL of 8 M urea, added to 7 mL of 1 M NaHCO3, and adjusted to pH 2.0 using 1 M HCl. Gastric juice was added at a 1:20 weight ratio and the mixture was incubated at 37 ⁰C for 1 h under static conditions. Gastric digestion was stopped by pH adjustment to 5.5 using 1 M NaHCO3. The pH was further adjusted to 7.5 using 1 M NaOH. Pancreatic juice was added at a1:10 ratio, and the mixture was incubated at 37 ⁰C for 2 h under static conditions. Pancreatic digestion was stopped by incubating the mixtures at 95 – 100 ⁰C for 10 min, followed by cooling and centrifugation at 8,000 rpm for 30 min at 4 ⁰C. The supernatant, which contained the protein hydrolysate, was immediately transferred into clean tubes.
Desalting and fractionation of protein hydrolysates
The supernatant was loaded onto a Supelclean LC-18 RP SPE column (Supelco, 5g/20 mL) conditioned with acetonitrile (ACN) and equilibrated with ultrapure water. The column was desalted by washing with water, and bound protein hydrolysates were eluted in sequence with 50 mL volumes of 20%, 35%, and 50% ACN containing 0.05% TFA; five 10-mL fractions were collected for each concentration of eluent. Fractions of the same percentage of ACN were pooled together, concentrated in vacuo, and lyophilized. The protein content was determined using BCA assay kit (Thermofisher Scientific).
HMG-CoA reductase Inhibitory assay
Inhibition of HMG-CoA reductase enzyme activity was assayed according to Villalobos et al. (2021) 35 using the assay kit CS1090, which is based on the decrease in the absorbance of NAPDH. Pravastatin (Sigma) was used as the reference standard treatment drug.
Tricine-SDS-PAGE
Tricine-SDS-PAGE was conducted according to Schägger (2006) 36. The samples were run under reducing conditions on a Mini-Protean tetra-cell (Bio-Rad) at an initial voltage of 30 V for 30 min, followed by an increase to 150 V for 90 min. The 1X anode and 2X cathode tank buffers were cooled throughout the run. The gel was then fixed, stained with Coomassie blue, and destained.
Animal model
Ethical considerations and housing conditions
All animal manipulation protocols were evaluated and approved by the UST IACUC (Code RC2017-950925). Acquisition of the animals and housing conditions were done as described by Villalobos et al. (2021) 35. Briefly, Sprague-Dawley rats were acquired and allowed to acclimatize for one week. They were housed in steel cages in a well-ventilated room maintained at a temperature of 22-25°C. The animals were allowed free access to purified drinking water throughout the duration of the study.
Cholesterol-lowering effect in vivo
Twenty Sprague-Dawley rats of mixed sexes, 6-8 weeks old and weighing 150 – 180 g, were acquired. Males were kept in separate cages from females. After acclimatization, the diet was changed ad libitum to a high cholesterol, high sugar diet consisting of 60% standard pellets, 15% lard, 10% egg yolk powder, and 15% sucrose for the next four weeks. After two weeks of a high-cholesterol diet, the rats were fasted overnight, and blood samples were drawn from the tail vein for analyses of total cholesterol (TC), high-density lipoprotein (HDL) and low-density lipoprotein + very low-density lipoprotein (LDL+VLDL) cholesterol.
The rats were then divided into four groups of five animals consisting of a mix of male and female animals, and age-matched per group (Table 1). TFPE was administered to one group and total protein hydrolysates (TPH) to another, while the positive control group was administered both acarbose and pravastatin. The untreated group served as a negative control. The treatments were administered daily for two weeks, via oral gavage in 0.5% saline vehicle. Animal BW was measured every 4 days for treatment dosing. TFPE was administered to Group 2 at 0.07 mg/kg BW, based on the dose for human consumption of lemongrass tea, which exerted significant but mild effects on renal parameters and blood pressure in human subjects 29,37. Group 3 was dosed at 0.05 mg/kg BW TPH. Acarbose and pravastatin were separately administered to Group 4 at the recommended doses of 40 mg/kg and 10 mg/kg, respectively 38,39. The untreated group received saline only. At the end of the treatment, the rats were fasted overnight and anesthetized with 0.1 mg/kg Zoletil. Blood samples were collected for TC, HDL, and LDL+VLDL analyses via intracardiac puncture, and the animals were sacrificed using Zoletil overdose. All blood samples were submitted for analysis of TC, HDL, and LDL+VLDL using the standard CHOD-PAP method for TC and direct measure-PEG for HDL, while LDL+VLDL was computed as the difference between TC and HDL 40.
Table 1: Animal groupings, treatments and dosage.
|
Animal grouping |
Treatment |
Dose (mg/kg BW) |
|
1 |
Untreated |
– |
|
2 |
TFPE |
0.07 |
|
3 |
TPH |
0.05 |
|
4 |
Acarbose+Pravastatin |
40+10 |
Statistical analysis
Differences between the change in cholesterol levels across treatments and cholesterol types were initially determined using two-way ANOVA with SPSS Software (IBM SPSS Statistics). Individual differences in cholesterol level changes were determined separately for treatment and cholesterol type using one-way ANOVA and Tukey’s HSD test in SPSS Software.
Results
Protein extract and hydrolysates
The yield of the TFPE was 0.03% (Table 2), consistent with other studies 41 which showed a relatively low yield of proteins from leaves in comparison to seeds, grains, or kernels 42. In vitro digestion of TFPE and subsequent fractionation on the SPE column yielded three fractions, with most proteins eluted as F1 (Figure 1). F1 accounted for 74% of TPH yield from digestion, while the yield of the other two fractions was approximately 1-2% only. Most of the hydrolysates were eluted with 20% ACN, indicating that the bulk of TPH is more polar relative to the 35% and 50% ACN fractions. F2 and F3 have decreasing polarity and are recovered in very small amounts. Tricine-SDS-PAGE revealed the presence of several bands with varying molecular weights in the fractions (Figure 2).
Table 2: Yield by weight of protein extraction and digestion
|
|
g ± SEM |
% Yield ± SEM |
|
Extraction of TFPE |
|
|
|
wt dried leaves |
621.43±53.29 |
|
|
wt TFPE |
0.202±0.057 |
0.03±0.01 |
|
Digestion of TFPE |
|
|
|
wt TFPE digested |
0.0578 ± 0.0010 |
|
|
wt 20% ACN (F1) |
0.0426 ± 0.0044 |
74.07 ± 8.69 |
|
wt 35% ACN (F2) |
≤ 0.0012 |
≤2 |
|
wt 50% ACN (F3) |
≤ 0.0010 |
≤1 |
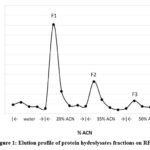 |
Figure 1: Elution profile of protein hydrolysates fractions on RP-SPE. |
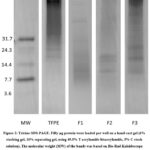 |
Figure 2: Tricine-SDS-PAGE. Fifty μg protein were loaded per well on a hand-cast gel (6% stacking gel, 16% separating gel, using 49.5% T acrylamide-bisacrylamide, 5% C stock solution). |
Inhibition of HMG-CoA reductase
The three hydrolyzate fractions at 0.5 ppm concentration showed inhibitory activities against human HMG-CoA reductase, in comparison with the pravastatin control. F1 exerted the lowest inhibition of 30.29%, F3 had the greatest at 48.69% inhibition, and pravastatin inhibited 78.10% of enzyme activity. IC50 values for pravastatin, F1, F2, and F3 were determined at 0.25, 1.14, 0.82 and 0.76 ppm, respectively (Table 3). Among the fractions, F3 had the lowest IC50, indicating that it was the most effective inhibitor. It was also the least polar and eluted last among the three fractions.
The specific activities of each fraction are listed in Table 3. F3 exhibited the highest specific activity and lowest IC50 value. To test for possible synergy, the three fractions were combined, and the resulting solution inhibited HMG-CoA reductase activity by 61%, which was higher than any of the individual fractions (Table 3). Also, a higher specific activity of 3,628% inhibition per μg protein was obtained, which was approximately 2.75-fold greater than the sum of the individual fractions. The increase in activity with the combination of the fractions increased the percentage of inhibition of HMG-CoA reductase. Although a significant increase in the inhibition of HMG-CoA reductase activity was observed, this result is at best preliminary, and more tests are needed to confirm if synergism among the components of the fractions occurs.
To determine if the hydrolysates exhibited cholesterol-lowering activity in vivo, Sprague-Dawley rats were initially fed a high-cholesterol, high-sugar diet for two weeks, then administered TPH for another two weeks simultaneously with the high-cholesterol diet. Table 4 shows the difference in serum TC, HDL, and LDL+VLDL levels before and after treatment, with TPH exhibiting a significant reduction (p < 0.05) in TC among the others. HDL levels decreased in all treatment groups, but the decrease was not significant. The effects of the treatments were clearly reflected in LDL+VLDL levels. Only the group treated with TPH showed a reduction in LDL+VLDL levels, while the other groups showed increased levels (Table 4). The untreated group showed the highest increase (p < 0.05), while the TFPE and Acarbose+Pravastatin groups experienced a moderate increase in LDL+VLDL levels.
Table 3: Inhibitory activity of fractions against HMG-CoA reductase
|
Fractions |
% Inhibition* ± SEM |
IC50 (ppm ± SEM) |
μg protein content# ± SEM |
% Inhibition per μg protein
|
|
Pravastatin |
78.10 ± 0.61 |
0.25 ± 0.02 |
|
|
|
F1 |
30.29 ± 2.26 |
1.14 ± 0.04 |
0.086 ± 0.001 |
350.63 ± 30.22 |
|
F2 |
46.25 ± 1.67 |
0.82 ± 0.07 |
0.103 ± 0.001 |
449.72 ± 16.25 |
|
F3 |
48.69 ± 4.44 |
0.76 ± 0.15 |
0.094 ± 0.001 |
515.38 ± 46.95 |
|
F1 + F2 + F3 |
61.31 ± 2.19 |
|
0.017 ± 0.001 |
3,628.01 ± 129.76 |
|
*the final concentration of the fractions and pravastatin was 0.5 ppm by weight, with corresponding protein content given in the table# |
||||
Table 4: Reduction in TC, HDL, and LDL+VLDL levels of Sprague-Dawley rats after treatment, mean mmol/L ± SEM.
|
Group |
Treatment |
TC |
HDL |
LDL+VLDL |
|
1 |
Untreated |
0.57 ± 0.16b,x |
0.72 ± 0.23x |
-0.21 ± 0.01c,y |
|
2 |
TFPE |
0.46 ± 0.11b,x |
0.63 ± 0.04x |
-0.09 ± 0.02b,y |
|
3 |
TPH |
0.86 ± 0.09a,x |
0.87 ± 0.07x |
0.01 ± 0.02a,y |
|
4 |
Acarbose+Pravastatin |
0.35 ± 0.07b,x |
0.45 ± 0.14x |
-0.11 ± 0.02b,y |
|
a-cSignificant differences between treatment type (P<0·05) x-ySignificant differences between cholesterol type (P<0·05) |
||||
Discussion
Recent studies have shown that bioactive peptides and protein hydrolysates produced by enzymatic digestion of proteins are HMG-CoA reductase inhibitors. We extracted tannin-free protein from lemongrass and determined its inhibitory effect on HMG-CoA reductase activity. Our results show that the hydrolysate fractions have good inhibitory activities on the HMG-CoA reductase enzyme, with IC50 values ranging from 0.76 to 1.14 ppm, versus that of pravastatin at 0.25 ppm. These values are also within the range of compounds that inhibit HMG-CoA reductase 9,11,43.
Potent HMG-CoA reductase inhibitor peptides released through simulated GI digestion could be of low molecular weight (MW) below 1 kDa, such as GGV, IVG, and VGVL from Amaranth protein 44, GCTLN from cowpea proteins 30, and VAWWMY from soy protein 45. There are also potent hypocholesterolemic peptides of higher MW, such as lunasin from soy with a 5.5 kDa MW 46, and a 6 kDa peptide from fermented food bekasam 47. In our study, the most active fraction, F3, could be long peptides or small proteins with MW ranging from 10-27 to kDa or higher, as visualized by SDS-PAGE. However, our results clearly show that peptides of low polarity in F3 exert greater HMG-CoA reductase inhibition than those of higher polarity in F2 or F1. Other authors have also emphasized that peptides with a high hydrophobicity index have the most potent cholesterol-lowering effect 48,49, which supports our results.
To support these in vitro studies, this study determined the potential of the fractions to reduce cholesterol in a rat model fed a hypercholesterolemic diet. The results showed a significant reduction in total cholesterol in rats administered TPH compared with controls. Although it seemed that only TPH treatment resulted in greater reduction of TC, while the untreated, TFPE, and control groups had the same degree of reduction, the results on the LDL+VLDL levels appear to further demonstrate the cholesterol-lowering effect of the treatments. Generally, statins influence the lowering of serum cholesterol levels by lowering LDL cholesterol but have less effect on increasing HDL levels 7. In this case, however, the control group containing pravastatin showed reduced LDL+VLDL levels, similar to the TFPE group, compared with the untreated group.
In conclusion, the protein hydrolysate fractions obtained from the digestion of lemongrass proteins were found to inhibit HMG-CoA reductase activity in vitro. Inhibition was observed in the individual fractions. Interestingly, when all fractions were combined, an approximately tenfold increase in inhibition per µg of protein was observed, which may imply cooperativity or synergism. Protein hydrolysates were fed to Sprague-Dawley rats, and the results showed a significant reduction in serum total and LDL+VLDL cholesterol levels in the test animals. Thus, lemongrass soluble proteins may serve as promising functional foods for the prevention of CVD risk. This study is the first to demonstrate the hypocholesterolemic effects of lemongrass protein hydrolysates . Currently, experiments are being conducted to identify bioactive peptides with the highest inhibitory activity against HMG-CoA reductase.
Acknowledgement
The authors thank Niel-Ju Angelle C. Cadiao and Mary Ann Julyn F. Catalan for their assistance with the animal study.
Conflicts of Interest
There are no conflicts to declare.
Funding Sources
This work is part of the dissertation of Mizpah C. Villalobos and is funded by Central Philippine University, Iloilo City Philippines; the Commission on Higher Education – Faculty Development Program (CHED-FDP), Diliman Quezon City Philippines, and the Department of Science and Technology – Philippine Council for Health Research and Development (DOST-PCHRD), Bicutan Metro Manila Philippines.
References
- Mensah GA, Roth GA, Fuster V. The global burden of cardiovascular diseases and risk factors: 2020 and beyond. J Am Coll Cardiol 2019;74(20):2529–2532; doi: 10.1016/j.jacc.2019.10.009.
CrossRef - Masic I, Alajbegovic J. The significance of the psychosocial factors influence in pathogenesis of cardiovascular disease. Int J Prev Med 2013;4(11):1223–1230.
- Rudd JHF, Davies JR, Weissberg PL. Atherosclerotic biology and epidemiology of disease. In: Topol EJ, Califf RM, Prystowsky EN, Thomas JD, Thompson PD, editors. Textbook of Cardiovascular Medicine. 3rd ed. Philadelphia, PA 19106: Lippincott Williams & Wilkins; 2007. p. 2–12.
- McNamara DJ. Cholesterol: Sources, absorption, function and metabolism Factors Determining Blood Levels. In: Encyclopedia of Human Nutrition. 2nd ed. Elsevier Ltd.; 2005. p. 379–385.
CrossRef - Burg JS, Espenshade PJ. Regulation of HMG-CoA reductase in mammals and yeast. Prog Lipid Res 2011; 50(4):403–410; doi: 10.1016/j.plipres.2011.07.002.
CrossRef - Istvan ES. Structural mechanism for statin inhibition of 3-hydroxy-3-methylglutaryl coenzyme A reductase. Am Heart J 2002;144(6 SUPPL.); doi:10.1067/mhj.2002.130300.
CrossRef - Pichandi S, Pasupathi P, Raoc YY, Farook J, Ambika A, Ponnusha BS, et al. The role of statin drugs in combating cardiovascular diseases. Int J Curr Sci Res 2011;1(2):47–56.
- Chen ZY, Jiao R, Ka YM. Cholesterol-lowering nutraceuticals and functional foods. J Agric Food Chem 2008;56(19):8761–8773; doi:10.1021/jf801566r.
CrossRef - Bok SH, Lee SH, Park YB, Bae KH, Son KH, Jeong TS, et al. Plasma and hepatic cholesterol and hepatic activities of 3-hydroxy-3- methyl-glutaryl-CoA reductase and acyl CoA: Cholesterol transferase are lower in rats fed citrus peel extract or a mixture of citrus bioflavonoids. J Nutr 1999;129(6):1182–1185.
CrossRef - Sashidhara K V., Singh SP, Srivastava A, Puri A, Chhonker YS, Bhatta RS, et al. Discovery of a new class of HMG-CoA reductase inhibitor from Polyalthia longifolia as potential lipid lowering agent. Eur J Med Chem 2011;46(10):5206–521; doi: 10.1016/j.ejmech.2011.08.012.
CrossRef - Ademosun AO, Oboh G, Passamonti S, Tramer F, Ziberna L, Boligon AA, et al. Phenolics from grapefruit peels inhibit HMG-CoA reductase and angiotensin-I converting enzyme and show antioxidative properties in endothelial EA.Hy 926 cells. Food Sci Hum Wellness 2015;4(2):80–85; doi: 10.1016/j.fshw.2015.05.002.
CrossRef - Erdmann K, Cheung BWY, Schröder H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J Nutr Biochem 2008;19(10):643–654; doi:10.1016/j.jnutbio.2007.11.010.
CrossRef - Nagaoka S. Structure–function properties of hypolipidemic peptides. J Food Biochem 2019;43(1):1–8; doi: 10.1111/jfbc.12539.
CrossRef - Ruiz JCR, Ancona DAB, Campos MRS. Proteínas y peptidos de origen vegetal en la reduccion de lipidos; potencial nutracéutico. Nutr Hosp 2014;29(4):776–784. doi:10.3305/nh.2014.29.4.7208.
- Rizzello CG, Tagliazucchi D, Babini E, Sefora Rutella G, Taneyo Saa DL, Gianotti A. Bioactive peptides from vegetable food matrices: Research trends and novel biotechnologies for synthesis and recovery. J Funct Foods 2016;27:549–569; doi: 10.1016/j.jff.2016.09.023.
CrossRef - Singh BP, Vij S, Hati S. Functional significance of bioactive peptides derived from soybean. Peptides 2014;54:171–179; doi: 10.1016/j.peptides.2014.01.022.
CrossRef - Al Disi SS, Anwar MA, Eid AH. Anti-hypertensive herbs and their mechanisms of action: Part I. Front Pharmacol 2016;6(JAN):1–24; doi: 10.3389/fphar.2015.00323.
CrossRef - Magotra S, Singh AP, Singh AP. A review on pharmacological activities of Cymbopogon citratus. Int J Pharm Drug Anal 2021;151–157; doi: 10.47957/ijpda.v9i2.465.
CrossRef - Oladeji OS, Adelowo FE, Ayodele DT, Odelade KA. Phytochemistry and pharmacological activities of Cymbopogon citratus: A review. Sci African 2019;6:e00137; doi: 10.1016/j.sciaf.2019.e00137.
CrossRef - Olorunnisola SK, Asiyanbi HT, Hammed AM, Simsek S. Biological properties of lemongrass: An overview. Int Food Res J 2014;21(2):455–462.
- Cheel J, Theoduloz C, Rodríguez J, Schmeda-Hirschmann G. Free radical scavengers and antioxidants from lemongrass (Cymbopogon citratus (DC.) Stapf.). J Agric Food Chem 2005;53(7):2511–2517; doi: 10.1021/jf0479766.
CrossRef - Rahim SM, Taha EM, Mubark ZM, Aziz SS, Simon KD, Mazlan AG. Protective effect of Cymbopogon citratus on hydrogen peroxide-induced oxidative stress in the reproductive system of male rats. Syst Biol Reprod Med 2013;59(6):329–336; doi: 10.3109/19396368.2013.827268.
CrossRef - Figueirinha A, Cruz MT, Francisco V, Lopes MC, Batista MT. Anti-inflammatory activity of Cymbopogon citratus leaf infusion in lipopolysaccharide-stimulated dendritic cells: Contribution of the polyphenols. J Med Food 2010;13(3):681–690; doi: 10.1089/jmf.2009.0115.
CrossRef - Costa CARA, Bidinotto LT, Takahira RK, Salvadori DMF, Barbisan LF, Costa M. Cholesterol reduction and lack of genotoxic or toxic effects in mice after repeated 21-day oral intake of lemongrass (Cymbopogon citratus) essential oil. Food Chem Toxicol 2011;49(9):2268–2272; doi: 10.1016/j.fct.2011.06.025.
CrossRef - Verma T, Sinha M, Bansal N, Yadav SR, Shah K, Chauhan NS. Plants used as antihypertensive. Nat Products Bioprospect 2021;11(2):155–184; doi: 10.1007/s13659-020-00281-x.
CrossRef - Garba HA, Mohammed A, Ibrahim MA, Shuaibu MN. Effect of lemongrass (Cymbopogon citratus Stapf) tea in a type 2 diabetes rat model. Clin Phytoscience 2020;6(1); doi: 10.1186/s40816-020-00167-y.
CrossRef - Dudai N, Weinstein Y, Krup M, Rabinski T, Ofir R. Citral is a new inducer of caspase-3 in tumor cell lines. Planta Med 2005;71(5):484–488; doi: 10.1055/s-2005-864146.
CrossRef - Thangam R, Sathuvan M, Poongodi A, Suresh V, Pazhanichamy K, Sivasubramanian S, et al. Activation of intrinsic apoptotic signaling pathway in cancer cells by Cymbopogon citratus polysaccharide fractions. Carbohydr Polym 2014;107(1):138–150; doi: 10.1016/j.carbpol.2014.02.039.
CrossRef - Ekpenyong C, Osim E. Changes in blood pressure indices in normotensive adults after the consumption of lemongrass tea. J Coast Life Med 2016;4(10):802–808; doi: 10.12980/jclm.4.2016J6-120.
CrossRef - Marques MR, Fontanari GG, Pimenta DC, Soares-Freitas RM, Arêas JAG. Proteolytic hydrolysis of cowpea proteins is able to release peptides with hypocholesterolemic activity. Food Res Int 2015;77:43–48; doi: 10.1016/j.foodres.2015.04.020.
CrossRef - Wang X, Chen H, Fu X, Li S, Wei J. A novel antioxidant and ACE inhibitory peptide from rice bran protein: Biochemical characterization and molecular docking study. LWT – Food Sci Technol 2017;75:93–99; doi: 10.1016/j.lwt.2016.08.047.
- Pivetta OH, Vaccaro MI, Maillie AJ. Decreased lipase activity in pure pancreatic juice and duodenal content from mutant mice with some alterations resembling cystic fibrosis. Life Sci 1981;28(20):2207–2213.
CrossRef - Hosseinzadeh H, Karimi G, Ameri M. Gastric irritation models in mice. BMC Pharmacol 2002;5:3–7; doi: 10.1186/1471-2210-2-21.
CrossRef - Capriotti AL, Caruso G, Cavaliere C, Samperi R, Ventura S, Zenezini Chiozzi R, et al. Identification of potential bioactive peptides generated by simulated gastrointestinal digestion of soybean seeds and soy milk proteins. J Food Compos Anal 2015;44(August):205–213; doi: 10.1016/j.jfca.2015.08.007.
CrossRef - Villalobos MC, Nicolas MG, Trinidad TP. Antihyperglycemic and cholesterol-lowering potential of dietary fibre from lemongrass (Cymbopogon citratus Stapf.). Mediterr J Nutr Metab 2021;14(4):453–67; http://dx.doi.org/10.3233/mnm-210568.
CrossRef - Schägger H. Tricine-SDS-PAGE. Nat Protoc 2006;1(1):16–22; doi:10.1038/nprot.2006.4.
CrossRef - Ekpenyong CE, Daniel NE, Antai AB. Effect of lemongrass tea consumption on estimated glomerular filtration rate and creatinine clearance rate. J Ren Nutr 2015;25(1):57–66; doi: 10.1053/j.jrn.2014.08.005.
CrossRef - Crespo MJ, Quidgley J. Simvastatin, atorvastatin, and pravastatin equally improve the hemodynamic status of diabetic rats. World J Diabetes 2015;6(10):1168; doi: 10.4239/wjd.v6.i10.1168.
CrossRef - Zhang Q, Xiao X, Li M, Li W, Yu M, Zhang H, et al. Acarbose reduces blood glucose by activating miR-10a-5p and miR-664 in diabetic rats. PLoS One 2013;8(11):1–9; doi:10.1371/journal.pone.0079697.
CrossRef - Warnick GR, Remaley AT. Measurement of cholesterol in plasma and other body fluids. Curr Atheroscler Rep 2001;3(5):404–411.
CrossRef - Yeoh HH, Wee YC. Leaf protein contents and nitrogen-to-protein conversion factors for 90 plant species. Food Chem 1994;49(3):245–250.
CrossRef - Day L. Proteins from land plants – Potential resources for human nutrition and food security. Trends Food Sci Technol 2013;32(1):25–42; doi: 10.1016/j.tifs.2013.05.005.
CrossRef - Li X, Liu C, Duan Z, Guo S. HMG-CoA reductase inhibitors from Monascus -fermented rice. J Chem 2013;2013; doi: 10.1155/2013/872056.
CrossRef - Soares RAM, Mendonça S, de Castro LÍA, Menezes ACCCC, Arêas JAG. Major peptides from Amaranth (Amaranthus cruentus) protein inhibit HMG-CoA reductase activity. Int J Mol Sci 2015;16(2):4150–4160; doi:10.3390/ijms16024150.
CrossRef - Nagaoka S, Nakamura YA, Shibata H, Kanamaru Y. Soystatin (VAWWMY), a novel bile acid-binding peptide, decreased micellar solubility and inhibited cholesterol absorption in rats. Biosci Biotechnol Biochem 2010;74(8):1738–1741; doi: 10.1271/bbb.100338.
CrossRef - Lule VK, Garg S, Pophaly SD, Hitesh, Tomar SK. Potential health benefits of lunasin: a multifaceted soy-derived bioactive peptide. J Food Sci 2015;80(3):C485–494; doi: 10.1111/1750-3841.12786.
CrossRef - Rinto, Dewanti R, Yasni S, Suhartono MT. Novel HMG-CoA Reductase inhibitor peptide from Lactobacillus acidophilus isolated from Indonesian fermented food bekasam. J Pharm Chem Biol Sci 2017;5(November):195–204. Available from: https://www.jpcbs.info/2017_5_3_05_Maggy.pdf.
- Lin SH, Chang DK, Chou MJ, Huang KJ, Shiuan D. Peptide inhibitors of human HMG-CoA reductase as potential hypocholesterolemia agents. Biochem Biophys Res Commun 2015;456(1):104–109; doi: 10.1016/j.bbrc.2014.11.042.
CrossRef - Zhong F, Zhang X, Ma J, Shoemaker CF. Fractionation and identification of a novel hypocholesterolemic peptide derived from soy protein Alcalase hydrolysates. Food Res Int 2007;40(6):756–762; doi:10.1016/j.foodres.2007.01.005.
CrossRef
List of Abbreviations
ACN – acetonitrile
BCA – bicinchoninic acid
BW – body weight
CHD – coronary heart disease
CVD – cardiovascular disease
GI – gastrointestinal
HDL – high-density lipoprotein
HMG-CoA – 3-hydroxy-3-methylglutaryl coenzyme A
IACUC – Institutional Animal Care and Use Committee
IC50 – Inhibitory activity, expressed as the amount of bioactive compound which inhibits 50% of the enzyme activity
LDL – low-density lipoprotein
RP-SPE – reverse-phase solid-phase extraction
TC – total cholesterol
TFPE – tannin-free
protein extract
TPH – total protein
hydrolysates
US-FDA – United States
Food and Drug Administration
UST – University
of Santo Tomas, Philippines
VLDL – very low-density lipoprotein

This work is licensed under a Creative Commons Attribution 4.0 International License.








