
Introduction
As a commodity chemical, acrylamide has numerous laboratory and industrial applications. After being cooked at high temperatures, many starchy foods naturally produce acrylamide through chemical reactions. Potato chips, French fries, grains (including breakfast cereals, toast, and cookies), coffee, and foods produced from grains are a few examples of foods with higher levels of acrylamide 1. When reducing, sugars (fructose or glucose) and the amino acid asparagine interact as part of the Maillard reaction, which is the mechanism that causes food to brown during roasting, frying, and baking; acrylamide is produced as a cooking process byproduct. Thus, elements such as cooking duration and temperature, moisture content, and the quantity of reducing asparagine and sugar in raw foods all have an impact on acrylamide levels in cooked food 2. Health risks to the general population based on an average exposure of 1 μg kg−1 body weight per day, increased to 4 μg kg−1 body weight per day for high consumers. Several epidemiological studies have performed an evaluation of the potential association with neurotoxicity as having the main toxic impact after occupational exposure. It is also toxic for reproduction, affecting male germ cells and being carcinogenic to a number of organs, including reproductive organs, gastrointestinal tract, kidneys, lungs, and brain 3,4.
Of any food evaluated, a sprout has an almost complete nutritional profile. Sprouts are full of vitamins, minerals, enzymes, and proteins. The ancient Egyptians and Chinese were aware of the nutritional benefits thousands of years ago, whereas seed germination and sprout production were used by the ancients. Sprouts are an organic food that improve seed quality through enhancing digestibility and decreasing the concentration of resistant starch and anti-nutritive chemicals 5.
Broccoli sprouts (Brassica oleracea L. var. italica) have small green leaves and short white stems that are connected to the seed. Broccoli sprouts are nutritious powerhouses, high in protein, fiber, vitamins (A and C), minerals (calcium and iron), and phytochemicals such as glucosinolates, polyphenols, and carotenoids. Sprouts have been linked to a number of biological qualities, including anti-diabetic, anticarcinogenic, anti-inflammatory, and antioxidant effects. Broccoli sprouts contain sulforaphane, a compound that could result in cancer cell apoptosis, or programed cell death, form new blood vessels that feed malignant cells, reduce inflammation, inhibit angiogenesis, reduce cancerous cell invasion and spread, lower susceptibility to cancer-causing substances, and increase certain detoxifying enzymes in the liver 6,7.
As a nutritious agricultural commodity, pea (Pisumsativum L) is part of the Leguminosae family. Pea sprouts have important nutrients that can be subdivided into soluble and insoluble fibers, proteins (lysine and tryptophan), complex carbohydrates, folate, vitamin B, minerals such as potassium, iron, calcium, but lower contents of sodium, cholesterol, and saturated fat 8. Pea proteins range between 244 and 275 g kg−1 and starch contents range between 440 and 462 g kg−1 on a dry basis with soaking, germination, fermentation, pasteurization, and including advanced techniques such as pulse electric field or ultrasonication as different processing methods, having demonstrated an excellent efficacy by increasing the nutritional benefits and techno-functional attributes of peas and its proteins 9, 10. Furthermore, pea sprouts include significant levels of antioxidant components like phenols, tocopherols, and carotenoids, which help the body to fight free radical damage that can result in disease. It has a number of health benefits, including lowering colon cancer prevalence, promoting type 2 diabetes, cardiovascular disease, and LDL-cholesterol. Pea sprouts naturally help diabetic individuals to monitor their blood sugar levels 11. Therefore, the objective of this study is to ascertain the chemical constituents of broccoli and pea sprouts and their effects on biological, biochemical and immunity parameters of rats intoxicated with acrylamide.
Material and Methods
Materials
99% pure acrylamide (C3H5ON) was purchased from Sigma (Diesenhofen, Germany), and other analytical grade chemicals and kits utilized in the analyses were purchased from Merck, Darmstadt, Germany.30 adult male albino Sprague Dawley rats (160± 5gm and 14-16 weeks old) were obtained from the medical research center of Jazan University, Jazan, Kingdom of Saudi Arabia. The basal diet comprised of 10% corn oil as a lipid source,10% casein as a protein source, 1% vitamin mixture, 0.5% choline chloride, 5% cellulose as a fiber source, 69.5% corn starch, and 4% salt mixture. Broccoli (Brassica oleracea L. var. italica) and pea (Pisumsativum L.) were purchased from Food Research Department, Field Crops Research Institute (FCRI), Agricultural Research Center (ARC), Giza, Egypt and were identified by Agricultural Research Station, Giza, Egypt.
Methods
Preparation of broccoli and pea sprouts
Broccoli and pea sprouts were soaked at a volume of 1/3 to 1 cup of peas and broccoli seeds in cold water for 8-12 hours then drained of soak water, rinsed and drained with cold water every 8-12 hours. The sprouts were kept in a low light location and were harvested on day 11 when short roots had sprouted on the majority of plants 12. The sprouts were heated in the air oven dryer (Fill Pack Industries, Faridabad) at 40 ͦC for 6h., milled and sieved using a 0.6 mm sieve to obtain the sprouts powder 13.
Determination of chemical constitutes total flavonoid, total phenols and antioxidants activity of broccoli and pea sprouts
Moisture, fat, protein and ash according to 14, dietary fiber was determined according to the methods of 15. Carbohydrate was ascertained by difference. Calorie values were calculated using values of 9 kcal/g for fat, 4 kcal/g for protein and carbohydrates as reported by16. Mineral contents were ascertained using an atomic absorption spectrophotometer (PerKin Elmer Instrument Model 2380, 63110 rodgau, Germany), as detailed by 17. Vitamin C was determined using colorimetric methods as per the method of 18. Beta carotene and Vitamin E contents were determined in accordance with 18 and 19, respectively. To determine total flavonoid, the aluminum chloride colorimetric method of 20was used. The quantity of phenols in each of the tested sprouts was ascertained with Folin-Ciocalteu reagent using the methods by 21. The ABTS radical scavenging activity of triplicate tested sprout samples was conducted following the method of 22. DPPH radical scavenging activity was assayed in accordance with 23. Estimating serum leptin concentration and serum insulin concentration were according to 24 and 25, respectively.
Induction toxic with acrylamide
During the experimental period, 25 rats received 2.5µg/kg body weight/ day to induce the acrylamide toxicity 26.
Experiment biological design
Preparation of the basic diet was conducted using the formula noted by 27. This involved using vitamin mixture components as preferred by 28. Formulation of the salt mixture was conducted in accordance with 29.
Table 1: Composition of basal and sprout diets
|
Ingredients |
Basal diet |
5% broccoli sprouts |
10% broccoli sprouts |
5% pea sprouts |
10% pea sprouts |
|
Casein |
10 |
10 |
10 |
10 |
10 |
|
Corn oil |
10 |
10 |
10 |
10 |
10 |
|
Cellulose |
5 |
5 |
5 |
5 |
5 |
|
Salt mixture |
4 |
4 |
4 |
4 |
4 |
|
Vitamin mixture |
1 |
1 |
1 |
1 |
1 |
|
Choline chloride |
0.5 |
0.5 |
0.5 |
0.5 |
0.5 |
|
Broccoli sprouts |
——- |
5 |
10 |
——— |
——– |
|
Pea sprouts |
—— |
———- |
———- |
5 |
10 |
|
Starch |
69.5 |
64.5 |
59.5 |
64.5 |
59.5 |
Ethical approval was acquired by the Scientific Research Standing Committee of Jazan University, (REC-44/08/583). Every biological experiment was carried out at the medical research center of Jazan University. Individually housed rats (n = 30 rats) in wire cages in a room were kept under healthy conditions and maintained at 25±2o C. For acclimatization, before the experiment began, each rat was fed on basal diet for one week. The rats were then put into six groups (5 rats each), the first of which was given a basal diet as a negative control group while the second was given a basal diet and 2.5μg acrylamide /kg bw/d to induce acrylamide toxicity as a positive control group. The third and the fourth groups were treated with 2.5μg acrylamide /kg bw/d and fed on a basal diet with broccoli sprouts at the levels of 5 and 10% while, rats in groups 5and 6 were treated with 2.5μg acrylamide /kg bw/d and fed on a basal diet with pea sprouts at the levels of 5 and 10% for 28 days as experimental period. The levels of different sprouts were added as replacement from the caloric content of added starch.
Biological evaluation
Diet was recorded daily, and body weight was recorded weekly. The body weight gain (BWG), feed efficiency ratio (FER), and some organ weights were obtained based on 30with the equations:
“Body weight gain = Final weight (g) – Initial weight (g)”
“Feed efficiency ratio (FER) = Body weight gain (g) / Feed intake (g)”
Blood sampling and organs
At the conclusion of each experiment, and after 12 hours fasting in every experimental group, blood samples were collected. This was done using the retro-orbital procedure using micro capillary glass heparinized tubes. Collection of blood samples were placed into clean, dry centrifuge tubes and left to clot for 30 minutes in a water bath ( 37o C ). To separate the serum, the blood was centrifuged at 3000rpm for ten minutes, and was aspirated into a clean cuvette tube and stored at – 20o C for analysis 31. Analysis of all serum samples were performed to determine glucose according to 32 method. Alanine aminotransferase (ALT) and aspartate aminotransaminase (AST) were measured according to 33 and 34, respectively. In addition, alkaline phosphatase (ALP) and total protein (Tp) were obtained based on 35and 36 methods, respectively. Globulin (Glb) and albumin (Alb) were obtained by 37. Uric acid, urea, and creatinine were ascertained in accordance with 38, 34 and 39. Measurement of total cholesterol and triglycerides was performed in accordance with 40 and 41, respectively, HDL, VLDL-c and LDL-c were conducted using the method advocated by 42 and 43, respectively. Glutathione peroxidase (GPX), superoxide dismutase (SOD), catalase (CAT), glutathione s-transferases (GSTs), total antioxidant capacity (TAC) and malondialdehyde (MDA) were measured according to 29, 44, 45, 46, 47, 48and 49, respectively. Monoaminooxidase (MAO), acetylcholinestrse (AChE), and creatine phosphokinase (CPK) were ascertained using the methods of 50, 51and 52, respectively. 53 was used to determine the immunoglobulin productions.
Simultaneously, the organs brain, liver, and pancreas were removed, washed in saline solution, wiped using filter paper, and weighed in accordance with 54.
Statistical analyses
SPSS software (Statistical Package for the Social Sciences, version 27, USA) was used to analyze the data, and was presented as mean ± SD, compared using a one-way ANOVA (analysis of variance) test followed by least significant difference (LSD) multiple range-test to determine significance of the intergroup. Significance was deemed to be a probability of p ≤0.05.
Results and Discussion
Chemical constituents of broccoli and pea sprouts
The proximate compositions of sprouts of broccoli and pea are shown in Table (2). The chemical composition reveals differences between the samples that are of statistical significance (p ≤0.05), according to the sprout types. The pea sprouts had significantly higher protein, fiber and ash than broccoli sprouts, while broccoli sprouts had high content in moisture, carbohydrates and energy content compared to pea content. In terms of fat content, there is no significant difference between the two sprouts. The increased metabolic activity during the early germination phases, which stimulate protein synthesis, are attributed to this rise in protein levels. In dicotyledonous plants, reserve proteins are enzymatically hydrolyzed to produce amino acids and peptides, which are then used to create new hydrolytic enzymes 55. The broccoli sprouts showed protein content from 16.16 to 34.4 g 100 g−1 of dry matter 56, and the obtained value is in the study range. In addition, broccoli sprouts displayed more protein contents than broccoli plant, with a value of 20.41 g 100 g−1 dry matter 57. Sprouts provide protein levels that are akin to high-protein diets. There was a reduction in total lipids seen during germination (Table 2). The aforementioned behavior is caused by a number of mechanisms in the lipid degradation metabolism by energy demand, which raises the respiratory rate55. The lipid content of broccoli sprouts has similarities to that reported by 58. The ash content of broccoli and pea sprouts were found to differ significantly (p 0.05).
The ash content increases with germination time; the 11-day sprouts had the highest concentration (7.21 g 100 g of dry matter), a 44% increase over 3-day sprouts. These outcomes were almost identical to 56 in broccoli inflorescences with an ash content of 7.95 g 100 g1. Most energy required to perform everyday tasks comes from consuming carbohydrates, which are primarily found in vegetables. Consuming fiber has been linked to a lower risk of diseases such as arteriosclerosis, hemorrhoids, colon cancer, and colitis 59. The crude fiber content of broccoli sprouts fell considerably (p ≤0.05) with respect to their germination period. Sprouts had the lowest value (4.91 of dry matter). This is attributable to the stage of the germination’s metabolic activation, which begins with the breakdown of reserve chemicals 55. Regarding the lipid content, growing sprouts in open fields resulted in high lipid content (4.65%). Similar trends have been found for carbohydrate content, which may be because during germination the carbohydrate reserves, particularly in legume seeds, are consumed as a source of energy to begin the germination process, as well as various other processes, occurring in the seeds. In addition, in open fields, because of direct sunlight, the young seedlings can photosynthesize, leading to a content increase in carbohydrate 60. However, 61 showed that peas had a low fat and carbohydrate; the decline is because they are used as an energy source during germination. The catabolism of lipids and carbohydrates to generate energy for seed germination may also contribute to the loss of the dry weight and sprouted seeds give more energy as compared to normal seeds. 62 reported that pea sprouts had high dietary fiber content and this increase was due to changes in the polysaccharides present in cell wallssuch as glucose, celluloseand mannose, suggesting that these changes were due to an increase in the plant’s cell structureduring germination and led to an increase in ash content.
From the same table, broccoli sprouts had significantly higher content from detected minerals compared to pea sprouts (on dry weight basis). Cruciferous vegetables are characterized by their mineral potential. Minerals play a vital part in a number of biological processes, which benefits both humans and plants. In the broccoli sprouts tested, the K and P concentrations were the highest, followed by that of Mg. 63. The increment in minerals content could be linked to the light exposure in the sprouting process and absorbing elements from media grown in open fields, as found by 64. Yet, 65 found that pea shoots had a higher content of phosphorous and potassium in comparison to other leafy green vegetables. The results confirmed that mineral contents were affected by environment and growing media.
Table 2: Chemical constituents of 100g broccoli sprouts or pea sprouts
|
Nutrient components |
Broccoli sprouts |
Pea sprouts |
|
Moisture |
10.4 a±1.23 |
8.43 b±1.02 |
|
Protein |
23.99 b±0.22 |
34.93 a ±6.22 |
|
Fat |
3.82 a ±0.03 |
4.04 a ±0.89 |
|
Dietary fiber |
16.86 b±1.65 |
18.23 a ±1.04 |
|
Ash |
6.72 b±0.98 |
8.54 a ±0.29 |
|
Carbohydrates |
38.21a ±2.86 |
25.83 b±6.96 |
|
Total calories |
283.18 a ±5.32 |
279.4 b±3.41 |
|
Iron (mg) |
11.89 a ±1.77 |
6.77 b±0.65 |
|
Phosphorous (mg) |
741.65 a ±9.64 |
206.55 b±5.32 |
|
Potassium (mg) |
898.06 a ±6.83 |
434.34 b±11.22 |
|
Magnesium(mg) |
537.89 a ±8.79 |
84.72 b±7.23 |
|
Sodium (mg) |
37.5 a ±1.65 |
24.63 b±3.29 |
|
Calcium (mg) |
228.07 a ±3.58 |
149.22 b±4.32 |
|
Zinc (mg) |
5.07 a ±0.67 |
1.34 b±0.07 |
|
Selenium(µg) |
9.12 a ±0.34 |
0.75 b±0.02 |
Values are means ± SD.
The values are expressed means ± SD. Significant differences were found in the means from the same column with different litters (p≤0.05).
Vitamins, total phenols, total flavonoid contents and antioxidants activity of broccoli and pea sprouts (per100g)
Table (3) presents the broccoli and green pea sprout vitamins. Broccoli sprouts had significantly higher content of tested vitamins compared to pea sprouts. Vitamin E was the highest vitamin, followed vitamin A while pea sprouts had vitamin A, which was the highest compared to the other tested vitamins. During sprouting (germination), a number of enzyme systems became active and led to significant changes in the nutrition of pulses. Vitamin C, which is essentially absent in dry legume seeds, experienced an increase after sprouting 65. However, 61 found that pea sprouts had a higher content of vitamin C, in contrast to other prepared leafy green veggies. The ascorbic acid in broccoli sprouts has been directly linked to the regulation of plant growth, especially in the early stages of embryo germination. As a result, sprout development, which includes vitamin C production reactivation, favors the vitamin C concentration in sprouts 66. Sprouts of broccoli are a great source of fiber, sulforaphane, vitamin C, and other minerals. Flu causing viruses are fought off by vitamin C, a potent natural antioxidant and immune system stimulator found in broccoli sprouts. It also has significant levels of another antioxidant, Vitamin A. It includes pro-vitamins such as carotene and zeaxanthin, which protect against macular degeneration of the retina and maintain skin integrity in the elderly 63.
In the same, data show the total phenols, total flavonoid content, and antioxidants activity. The total phenols content was found to have a significant change (p≤0.05) between broccoli and pea sprouts. The pea sprouts showed high content of total phenols compared to broccoli sprouts, while the broccoli sprouts significantly increased in total flavonoid contents compared to pea sprouts. These results are similar to 67 with an improved total phenols content in the older sprouts. The concentration differences of these plant metabolites fluctuate with variety, sample treatment, development stage, quantification method, and extraction process 68. Broccoli sprouts total flavonoid content experienced a significant increase with the germination time, with the old sprouts increasing above 55% in flavonoid content 69. The tissue age, variety, and environmental factors such as temperature and light could be the cause of variations in total flavonoid concentration between broccoli sprouts and the previously reported values. For example, the production of anthocyanins is frequently accelerated in aged tissues, and its quantity rises with light 70. Pea seeds had a higher concentration of total phenols during germination than non-sprouted seeds, reaching day 6 maximum concentrations for bean sprouts of 70.04% higher than in the non-sprouted seeds. The pea’s maximum TP concentration was attained on day 7, increasing by 55.85% from non-sprouted seeds, and phenolic compound synthesis was a reaction to the pea’s germination71 found an increase in total flavonoid concentrations in the pea seed 61. The quantity of TP of the peas and their sprouts is more than that of fruits such as lychee, papaya, banana, pineapple and their by-products, demonstrating their capacity as a bioactive compound source 61. The DPPH procedure is a fast, simple, and appropriate method (in vitro) for evaluating the antioxidant capacities of vegetables and fruits 72. The quantification of total phenols and total flavonoids revealed similar behavior in the results of the DPPH radical scavenging capacity of pea and broccoli sprout (Table 3). When comparing the DPPH radical scavenging ability of pea sprouts to broccoli sprouts, there was a progressive rise (p >0.05). The greatest antioxidant levels were in sprouts, and multiple studies have shown a connection between total phenols content, vitamins, and antioxidant activity 73,74. Ferric reducing antioxidant power (FRAP) assay, ABTS radical cation assay, or oxygen radical absorbance capacity (ORAC) assay can confirm the antioxidant properties of broccoli and pea sprouts, which is closely related to phenols compounds, and there were significant correlations between total phenols and ABTS 75,65.
Table 3: Vitamins, total phenols, total flavonoid contents and antioxidants activity of broccoli and pea sprouts (per100g)
|
Variables |
Broccoli |
Pea |
|
Vit.C (mg) |
51.62 a ±8.54 |
32.65 b±1.34 |
|
Vit.E (IU) |
749.88 a ±10.43 |
6.65 b±0.82 |
|
Vit. A (IU) |
210.45 a ±9.76 |
199.87 b±5.72 |
|
Total phenols μgGAE/g |
70.87 b ±4.87 |
830.65 a ± 10.92 |
|
Total flavonoid μgGAE/g |
110.64 a ±6.83 |
67.34 b± 0.99 |
|
DPPH (%) |
39.76 b ± 7.93 |
79.65a ± 5.32 |
|
ABTS (%) |
67.65a± 1.03 |
49.65 b ± 2.04 |
Values are means ± SD.
The values are expressed means ± SD. Significant differences were found in the means from the same column with different litters (p≤0.05).
The Table (4) results illustrate the effect of tested sprout levels on body weight gain (BWG), feed efficiency ratio (FER), and feed intake (FI) forrats treated by acrylamide as toxic material. It is apparent that the normal rats fed with a basal diet recorded normal BWG, FI g/28 day and FER while the positive control group was significantly increased (p ≤0.05) when compared to the other groups. The greatest effects of the FI, BWG and FER in rats treated with tested toxic material was recorded for pea sprouts groups followed by broccoli sprouts groups and fed on diet containing 10% from tested sprouts seemed to reveal a high gradual decreasing of BWG, FI and FER followed by 5%. No significance was found between the sixth group and negative group. ACR is contained within a great deal of foods and is produced when root vegetables or potatoes that contain the amino acid asparagine are cooked at a high temperature with foods that contain a high concentration of carbohydrates 76.
Acrylamide induces obesity and has been linked to abdominal obesity and weight gain in humans. However, there is an absence of clarity on the molecular mechanism whereby acrylamide can induce adipocyte differentiation and obesity through lipid metabolism 77. Exposure to acrylamide may have an effect on thyroid function, and it is possible that the altered thyroid influences body weight. However, exposure to acrylamide has been linked to hypothyroidism, which results in weight gain instead of weight loss 78. Sprouts are high in fiber and low in calories, as well as an incredible source of protein. These factors make it an excellent weight-loss food. They help improve satiety and add to daily protein requirements. Broccoli sprouts are rich in a range of nutrients, including fiber, vitamin A, vitamin C, and sulforaphane. Sulforaphane results in 20% fat loss by altering gut bacteria and increasing mitochondria in fat in mice 69. Pea sprouts are an excellent source of micronutrients, they provide protein which promotes satiety, low fat and fiber as well as providing vitamins C, A, K, and other vital vitamins and minerals 71.
Table 4: Body weight gain (BWG), feed intake (FI) and feed efficiency ratio (FER) of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts
|
Groups |
Parameters |
||
|
BWG (g/28d) Mean ±SD |
FI (g/day) Mean ±SD |
FER (g/rat/day) Mean ±SD |
|
|
Negative control (G1) |
40.04d± 2.67 |
15.22c ±1.12 |
0.094 e±0.001 |
|
Positive control(G2) |
63.32a± 0.91 |
20.22a ±0.71 |
0.112a±0.002 |
|
5% broccoli sprouts (G3) |
55.90b±1.53 |
18.15b ± 0.99 |
0.110b±0.001 |
|
10% broccoli sprouts (G4) |
47.40c± 0.64 |
16.42c ± 0.76 |
0.103d±0.003 |
|
5% pea sprouts (G5) |
50.48c±1.89 |
17.17b± 1.04 |
0.105c±0.004 |
|
10% pea sprouts (G6) |
41.68 d ±2.05 |
15.67 c ±0.78 |
0.095 e ±0.001 |
|
|
3.77 |
1.63 |
0.001 |
The values are expressed means ± SD. Significant differences were found in the means from the same column with different litters (p≤0.05).
The results in figure (1) show the effects of levels of pea and broccoli sprouts in rats treated with acrylamide on liver, kidney, brain and pancreas weights. The results showed that the weight of organs significantly increased (p≤0.05) in the positive control group in comparison to other groups. The best level was 10% which s had a high effect on organs weight, but they remained significantly higher (p≤0.05) than the negative control group, except kidney weight. The obtained results were matched with the results of 77 who found that acrylamide at the dose 2 μg kg/ body weight / day per 35 days caused a significant increase in relative brain and liver weight. Sprouts are rich in many important nutrients which, depending on the type of sprout, contain high levels of carbohydrates, protein, dietary fibers, folate, magnesium, phosphorus, and vitamin K. They also possess antioxidant and antimutagenic activities which protect the internal organs from free radical and toxic material 69. Germination is a simple and inexpensive way to improve nutritive value. A diet that is high in broccoli sprouts offers an excellent source of vitamin C, indole-3-carbinol, flavonoids, sulforaphane, and minerals and might lower the risk of organ diseases, especially cancer 57. Peas contain bioactive compounds including micro-nutrients, macronutrients, and phytochemicals. Phytochemicals, usually polyphenols, such as flavonoids and phenolics compounds, are known to provide anti-oxidative and anti-inflammatory properties in internal organs 71.
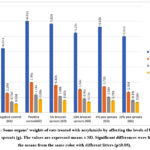 |
Figure 1: Some organs’ weights of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts (g).
|
The results in table (5) show the effects of tested sprouts levels on serum glucose, insulin and leptin hormones. As revealed in the table, group 6 had the best glucose level and tested hormones; this group was toxic with acrylamide and fed on 10% pea sprouts followed by the same level of broccoli sprouts. There were significant(p≤0.05) changes between both controls and different tested groups.79 discovered that there is a large positive dose-related link between acrylamide exposure and an increase in fasting plasma glucose77 indicated that insulin resistance was related to acrylamide exposure and a decrease in insulin level. Male rats given acrylamide at doses of 25 mg/kg b.w./d and 50 mg/kg b.w./d reduced their beta cells by 15% and 25%, impairing normal blood levels of glucose and insulin 80. The concentration of plasma leptin increases proportional to body fat mass and regulates food intakes and energy outlay by using brown fat for energy and excessive hunger to maintain body fat stores. Leptin is known to inhibit insulin secretion by PI3K-dependent activating phosphodiesterase 3B (PDE3B) and by decreasing the level of cAMP, therefore inhibiting the protein kinase A (PKA) pathway and is a regulator of Ca2+ channels and exocytosis 81,82. Sprouts can help to control blood sugar levels because they contain lower levels of carbohydrates, which can help to regulate insulin levels. Pea sprouts can lower blood glucose levels. The fiber and protein content of peas slows digestion and smoothens the body’s blood sugar levels 83. Broccoli has a very low Glycemic Index and can also help regulate blood sugar levels. Furthermore, broccoli contains a lot of fiber, which can help regulate appetite and maintain stable blood sugar levels. Research has shown that sulforaphane-rich broccoli extract has potent anti-diabetic benefits, lowering blood sugar and oxidative stress indicators, and improving insulin sensitivity 84.
Table 5: Serum glucose, insulin and leptin hormones of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts
|
Groups |
Parameters |
||
|
Glucose mg/dl Mean ±SD |
Insulin µU/mL Mean ±SD |
Leptin ng/mL Mean ±SD |
|
|
Negative control (G1) |
104.66f± 5.94 |
17.43a ±1.11 |
12.34 f±1.02 |
|
Positive control(G2) |
206.54a± 7.04 |
6.18f±0.68 |
25.08a ±0.99 |
|
5% broccoli sprouts (G3) |
185.09b±4.51 |
7.80e±0.94 |
22.18b ± 1.04 |
|
10% broccoli sprouts (G4) |
157.12d± 6.21 |
10.87c±1.21 |
18.50d ± 0.87 |
|
5% pea sprouts (G5) |
175.33c±5.62 |
9.05d±0.93 |
20.40c ± 1.28 |
|
10% pea sprouts (G6) |
135.29 e±4.81 |
12.07 b±1.04 |
16.51 e±0.08 |
|
LSD |
7.65 |
1.24 |
1.77 |
The values are expressed means ± SD. There is a significant difference in means from the same column with different litters (p≤0.05).
Figures (2 ,3) shows that significant increases occurred in the concentration of the liver enzymes (AST, ALT and ALP) and total protein which equal the sum of albumin and globulin in the blood serum in the positive control group in comparison to the negative control group. The groups which were toxic with AA and treated with tested sprouts showed significant improvement in tested parameters in comparison to positive control group but were still higher than the negative control group. The 10% level from tested sprouts had the greatest effect followed by 5%, and the pea sprouts recorded a higher effect than the broccoli sprouts. Aspartate aminotransferase (AST), Alanine aminotransferase (ALT), and alkaline phosphatase (ALP) are enzymes, which when altered signify liver damage. Acrylamide causes liver damage and increases the levels of those enzymes85. Sprouts contain phenols and vitamins which play a vital role as a strong antioxidant in scavenging free radicals, reducing cell toxicity and death, and are also anticarcinogenic. Solforavan compound extracted from broccoli decreases liver damage by causing a decrease in the amounts of AST, ALT, and ALP levels and to activate Nrf2/Keap pathway 86. Peas have low-fat-rich protein, high fiber and high content of antioxidant components mainly including phenols, tocopherols, and carotenoids, which significantly alleviate oxidative stress in the liver 87.
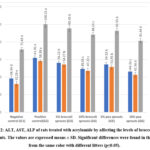 |
Figure 2: ALT, AST, ALP of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts. |
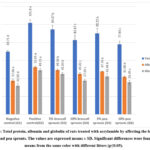 |
Figure 3: Total protein, albumin and globulin of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts. |
The results in table (6) show that the positive control group’s levels of uric acid, urea, and creatinine were significantly higher (p >0.05) than those of the negative control group. When compared to the toxic positive control group, all toxic rats with AA who were fed a diet that contained 5% and 10% broccoli and pea sprouts showed significantly lower kidney function levels. The group 6 toxic rats fed 10% pea sprouts received the best care. The 10% of examined sprouts had no significant difference between the groups. The findings showed that oral exposure to acrylamide through drinking water or gavages causes chronic liver disease, hepatic insufficiency, and kidney damage, leading to primary immunodeficiency and immune system activation after the exposure88. The kidneys of the acrylamide-treated groups exhibited glomerular tuft degradation and lymphocyte infiltration. The epithelial lining of the renal tubules had degenerative alterations, which were followed by cell rupture, necrosis, and interstitial blood vessel congestion. The renal tubules also lost their brush boundaries and became vacuolated 89. Compounds found in broccoli could help to treat or halt the course of CKD. Numerous phytochemicals can increase NRF2 and offer oxidative protection. Glucosinolates(-thioglucoside N-hydroxysulfates), such as the glucoraphanin found in broccoli90. Treatment with pea sprouts can lower blood pressure and provide protection against hypertensive kidney injury by boosting anti-inflammatory and antioxidant defense responses and regulating the renin-angiotensin system 87. Uric acid, urea, and creatinine levels in the serum of the administered rat were considerably lower (P<0.05) than in the control group. Reductions in the concentrations of uric acid, urea, and creatinine were discovered to be dose-dependent.
Table 6: Affecting the levels of broccoli and pea sprouts on creatinine, urea, and uric acid of rats treated with acrylamide
|
Group |
Parameters |
||
|
Creatinine (mg/dl) |
Urea (mg/dl) |
Uric acid (mg/dl) |
|
|
Negative control (G1) |
0.73e±0.06 |
18.34d±2.05 |
1.23d±0.18 |
|
Positive control(G2) |
1.16a±0.04 |
29.05a±2.31 |
2.91a±0.29 |
|
5% broccoli sprouts (G3) |
1.06b±0.01 |
25.80b±2.12 |
2.45b±0.01 |
|
10% broccoli sprouts (G4) |
0.98c±0.002 |
20.22d±2.12 |
1.96c±0.18 |
|
5% pea sprouts (G5) |
0.97c±0.08 |
24.75c±1.23 |
2.21b±0.21 |
|
10% pea sprouts (G6) |
0.88d±0.02 |
19.05d±2.43 |
1.71c±0.061 |
|
LSD |
0.090 |
2.89 |
0.28 |
The values are expressed means ± SD. Significant differences were found in the means from the same column with different litters (p≤0.05).
The values are expressed means ± SD. Significant differences were found in the means from the same column with different litters (p≤0.05).
Table (7) illustrates the mean values of serum lipid profile concentration in both of controls and toxic groups with AA and treated with levels of tested sprouts. There were significant (p≤0.05) increases of TC, TG, (LDL-c), (VLDL-c) and aside from a significant (p≤0.05) reduction of (HDL-c) of toxic control group with AA in comparison to negative control group rats. Moreover, every toxic rat fed on diet that contained 5% and10% tested sprouts revealed marked decreases of TC, TG, (LDL-c) and (VLDL-c) as well dramatic increases of (HDL-c). The best results were for group 6 (rats fed on 10% pea sprouts) where it had a potent effect. No significant differences were found between groups fed on 10% broccoli sprouts and 5% pea sprouts for TC, TG, (LDL-c) and (VLDL-c). The data obtained by91shows how AA affected the lipid profile in the blood, significantly raising TC and TG levels while significantly reducing HDL-C levels. The involvement of AA in lipid peroxidation can therefore account for the findings. High plasma oxidative stress indicators were negatively connected with low HDL and favorably correlated with elevated plasma triglycerides.
By inhibiting lipoprotein lipase and the reduction in VLDL clearance in adipocytes caused by impaired insulin signaling, AA exposure could result in atherogenic dyslipidemia. Additionally, the amount of free fatty acids (FFA) in the liver is increased, which acts as a substrate for the production of TG and causes hypertriacyglycemia. The formation of Apo B, the primary apoprotein of VLDL particles, is also stabilized by the FFA92, 93. Numerous trials have found that consuming sprouts decreases cholesterol in those with diabetes or obesity. One study found a decrease in triglycerides and “bad” LDL cholesterol and an increase in “good” HDL cholesterol.94 shows that broccoli sprouts could make improvements to lipid profiles and blood glucose. Plasma LDL-C was also significantly down regulated with intake of high glucoraphanin broccoli. According to 95, consumption of broccoli sprouts can reduce cardiovascular diseases and enhance lipid-related metabolic markers. 96 discovered that, compared to nonfermented pea, fermented pea significantly reduced the serum levels of TC, TG, and LDL in mice. This could be because aminobutyric acid found in fermented peas helps hypercholesterolemic mice by lowering their lipid levels. Additionally, it contains lectins, oligosaccharides, folate, selenium, zinc, saponins, and selenium, all of which reduce the oxidative stress that is strongly linked to hyperlipidemia97.
Table 7: Lipid profile of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts.
|
Group |
Parameters |
||||
|
Triglycerides (mg/dl) |
Total cholesterol (mg/dl) |
HDL-c (mg/dl) |
LDL-c (mg/dl) |
VLDL-c (mg/dl) |
|
|
Negative control (G1) |
97.61 e±4.05 |
99.68 e±3.12 |
50.74a ±2.55 |
29.42e ±2.58 |
19.52d ±0.45 |
|
Positive control(G2) |
129.4 a ±5.03 |
137.14 a ±3.11 |
39.62c ±2.34 |
71.64a ±1.82 |
25.88a ±0.55 |
|
5% broccoli sprouts (G3) |
125.75 b ±3.65 |
134.3 b±2.33 |
44.31b ±2.35 |
64.84b ±2.93 |
25.15a ±0.25 |
|
10% broccoli sprouts (G4) |
118.53 c±3.02 |
126.5 c±1.78 |
48.46a ±2.64 |
54.33c ±1.51 |
23.71b ±0.32 |
|
5% pea sprouts (G5) |
121.4 c±3.85 |
127.58 c±2.89 |
45.62b ±2.55 |
57.68c ±3.88 |
24.28b ±0.13 |
|
10% pea sprouts (G6) |
111.32 d±2.65 |
122.53 d±2.19 |
49.63a ±1.89 |
50.64d ±2.78 |
22.26c ±0.13 |
|
LSD |
6.75 |
3.32 |
2.67 |
3.91 |
1.45 |
The values are expressed means ± SD. There is a significant difference in the means from the same column with different litters (p≤0.05).
Table (8) shows that the positive control group’s activating levels of GPX, SOD, and CAT were considerably lower (p ≤0.05) than those of the negative control group. All rats given acrylamide treatment and diets containing 5 and 10% broccoli and pea sprouts had steady increases in antioxidant levels that were considerably (p >0.05) higher than those of the positive control group. Group 6 received the best therapy, which raised the antioxidant enzymes. The table also shows that, when positive controls were compared to negative controls, there were significant declines in the levels of GST and TAC as well as significant increases in MDA, but all toxic rats fed diets containing the tested sprouts experienced a significant improvement in GST levels98.Tested sprouts have a high antioxidant capacity and are rich in polyphenols, which impact upon antioxidant enzyme activities in vivo, for example an increase in SOD and GSH activities and a decrease in MDA levels 99.Broccoli sprouts have high antioxidant effects in human cells as a result of several protective compounds. They contain 10-100x more enzymes than fruits and vegetables when eaten without germination. They are rich in glucoraphanin, which is changed to another compound called sulforaphane during digestion. This chemical has antioxidant effects which might help prevent cancer and liver problems 100.Pea sprouts contain high levels of flavonoids and phenolic compounds, which are essential in, hepatic impairments, reducing inflammation in hepatic cells apoptosis, and structural injury in the liver while also inhibiting both types of structural injury via degradation of oxidative stress, DNA damage to the liver, and malondialdehyde (MDA), which is linked to liver damage caused by acrylamide 101, 102.
Table 8: Glutathione peroxidase (GPX), superoxide dismutase (SOD), catalase (CAT), glutathione S-transferases (GST), total antioxidant capacity (TAC) and malondialdehyde (MDA)of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts
|
Group |
Parameters |
|||||
|
GPX (ng/dl) |
SOD (U/L) |
CAT (mmoL/L) |
GST (mmoL/L) |
TAC (nmoL/L |
MDA (nmoL/L) |
|
|
Negative control (G1) |
83.73a±3.21 |
50.65a±2.9 |
73.04a±5.67 |
31.97a±2.49 |
1.63a±0.09 |
13.97e±1.88 |
|
Positive control(G2) |
58.43d±3.10 |
32.05d±4.19 |
42.83e±3.41 |
20.20d±4.08 |
0.83e±0.05 |
35.21a±4.05 |
|
5% broccoli sprouts (G3) |
63.54c±3.66 |
39.28c±3.52 |
48.09 d ±4.22 |
23.11c±2.06 |
0.99d±0.14 |
30.78b±2.74 |
|
10% broccoli sprouts (G4) |
70.89b±3.14 |
44.77b±3.21 |
56.38c±6.17 |
28.16b±1.73 |
1.33b±0.25 |
25.58c±4.22 |
|
5% pea sprouts (G5) |
65.48c±3.78 |
41.74b±3.82 |
52.15c±6.54 |
25.84b±1.22 |
1.30c±0.02 |
27.57b±3.44 |
|
10% pea sprouts (G6) |
73.16b±2.86 |
47.88a±2.43 |
61.55b±6.53 |
30.54a±2.67 |
1.38b±0.002 |
20.67d±2.45 |
|
LSD |
4.21 |
3.86 |
4.76 |
2.89 |
0.07 |
3.85 |
The values are expressed means ± SD. There was a significant difference in the means from the same column with different litters (p≤0.05).
Table (9) shows alterations to the serum parameters, including brain enzymes. A significant decrease was found in the activities of MAO and AChE in the positive control group, while CPK activities had a significant increase in the same group compared to the negative control group and the groups fed on 5% and 10% tested sprouts. The pea sprouts had a significant effect compared to broccoli sprouts. The levels of 10% of tested sprouts recorded a high effect compared to 5%. There was no significant variation between the 10% pea sprouts group and negative control group. Acrylamide interacts with various biochemical processes such as acetylcholine metabolism and therefore can act as a potential etiopathogenic cofactor. It causes AChE inhibition and decreased MAO activity, which may represent the mode of action by which AA contributes to pathological processes inducing neurotoxicity 98. Sprouts have high levels of omega-3 fatty acids and support neuronal health and decrease the risks of heart attack and stroke. Pea and broccoli sprouts have high levels of magnesium and calcium, which support bone and muscle health. Magnesium is crucial for vessel and brain health. Pea sprouts are an excellent source of dietary fiber, vitamin A, C and E, and potassium. Glycosylated quercetin derivatives are the primary flavonoid compounds of pea sproutsand protect the brain from damage and keep its enzymes within normal range. Broccoli sprouts are packed with vitamin K which can help limit the extent of brain neuron damage and help delay or prevent the onset of Alzheimer’s disease 100, 96.
Table 9: The brain enzymes of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts.
|
Group |
Parameters |
||
|
MAO U/ml |
AChE U/ml |
CPK U/l |
|
|
Negative control (G1) |
21.03 a ± 1.56 |
6.48 a ± ±1.08 |
22.57 d ± 2.84 |
|
Positive control(G2) |
10.64 d ± 1.93 |
1.22 e ± 0.84 |
34.06 a ± 2.89 |
|
5% broccoli sprouts (G3) |
13.67 c ± 1.17 |
2.79 d± 0.06 |
30.19 b ±2.54 |
|
10% broccoli sprouts (G4) |
17.98 b ± 2.66 |
4.59 c ± 0.98 |
26.34 c ± 2.81 |
|
5% pea sprouts (G5) |
16.59 b ± 1.05 |
3.01 d± 0.76 |
27.88 b ± 2.61 |
|
10% pea sprouts (G6) |
20.99 a ± 4.76 |
5.31 b ± 0.79 |
23.07 d ± 2.61 |
|
LSD |
2.92 |
0.99 |
2.88 |
The values are expressed means ± SD. There was a significant difference in the means from the same column with different litters (p≤0.05).
IGA, IGG and IGM of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts
Table (10) shows higher values in IGA, IGG and IGM for the negative control in comparison to the positive one. Significant differences were found in the mean values of tested parameters across the treatment groups (p ≤ 0.05). Highest values of tested parameters were found in the 10% pea sprouts group. Sprouts contain phenol compounds which are vital in defense responses, including anti-inflammatory, anti-aging, antioxidant and anti-proliferative activities. Broccoli and pea sprouts have higher vitamin C content, making them potent stimulants for the body’s white blood cells in order to fight disease and infection, thus, helping to build immunity. Also, they have a high number of phenols which act as an antioxidant by reacting with free radicals by transfer of a single electron, hydrogen atom transfer, chelation of transition metals, and sequential proton loss electron transfer. They also contain flavonoids which inhibit a number of enzymes such as phosphodiesterase, xanthine oxidase, aldose reductase and regulate different hormones such as thyroid hormones, estrogens, and androgens 103, 104.
Table 10: IGA, IGG and IGM of rats treated with acrylamide by affecting the levels of broccoli and pea sprouts
|
Groups |
Parameters |
||
|
IGA (mg/dl) |
IGG (mg/dl) |
IGM (mg/dl) |
|
|
Negative control (G1) |
140.44±9.72a |
610.3±7.92a |
150.63±9.08a |
|
Positive control(G2) |
80.73±5.82e |
445.74±9.06f |
70.37±7.69f |
|
5% broccoli sprouts (G3) |
91.95±4.89d |
467.93±8.34e |
79.75±5.91e |
|
10% broccoli sprouts (G4) |
102.05±7.44c |
507.65±10.04c |
95.91±5.67c |
|
5% pea sprouts (G5) |
97.34±10.22c |
487.87±10.94d |
86.97±6.48d |
|
10% pea sprouts (G6) |
116.98±9.45b |
527.67±8.99b |
107.67±5.48b |
|
LSD |
6.89 |
10.87 |
5.32 |
The values are expressed means ± SD. There are significant differences in the means from the same column with different litters (p≤0.05).
Conclusion
Consuming broccoli sprouts and pea sprouts is an economical, intelligent, and simple option to eat healthily. The broccoli and pea sprouts analyzed in this research are low in fat and rich in protein. The sprouts also have high contents of flavonoids, polyphenols, a high DPPH radical scavenging capacity and ABTS. The results indicate the benefit of tested sprouts consumption for anti-inflammatory activity, improved brain enzymes, decreased biological parameters (body weight gain, feed intake and some organs weight) and decreased the levels of tested biochemical parameters (fasting blood glucose, liver enzymes, kidney functions, lipid profile and MDA).
Conflict of Interest
The authors declare that there is no actual or potential conflict of interest.
Funding Sources
This work was supported by the Deanship of Scientific Research, King Faisal University, Saudi Arabia [GRANT 4700].
References
- Stadler, R. H., Blank, I., Varga, N., Robert, F., Hau, J., Guy, P. A., …&Riediker, S. (2002). Acrylamide from Maillard reaction products. Nature, 419(6906), 449-450.
CrossRef - Nematollahi, A., Meybodi, N. M., &Khaneghah, A. M. (2021). An overview of the combination of emerging technologies with conventional methods to reduce acrylamide in different food products: Perspectives and future challenges. Food Control, 127, 108144.
CrossRef - Erkekoğlu, P., &Baydar, T. (2010). Toxicity of acrylamide and evaluation of its exposure in baby foods. Nutrition research reviews, 23(2), 323-333.
CrossRef - Hogervorst, J. G., Schouten, L. J., Konings, E. J., Goldbohm, R. A., & van den Brandt, P. A. (2009). Dietary acrylamide intake and brain cancer risk. Cancer Epidemiology Biomarkers & Prevention, 18(5), 1663-1666.
CrossRef - Abdallah, M. M. F., Ghanem, K. M., Abo El-Naga, M. M., El-Sheshtawy, A. A., &Abd El-Gany, T. S. A. (2017). Effect of wheat flour supplemented with some seed sprouts flours on cake qualities. J. of Biolo. Chem. and Environ. Sci, 12(3), 113-130.
- Idowu, A. T., Olatunde, O. O., Adekoya, A. E., &Idowu, S. (2020). Germination: An alternative source to promote phytonutrients in edible seeds. Food Quality and Safety, 4(3), 129-133.
CrossRef - Le, T. N., Chiu, C. H., & Hsieh, P. C. (2020). Bioactive compounds and bioactivities of Brassica oleracea L. var. italica sprouts and microgreens: An updated overview from a nutraceutical perspective. Plants, 9(8), 946.
CrossRef - Robinson, G. H. J., Balk, J., &Domoney, C. (2019). Improving pulse crops as a source of protein, starch and micronutrients. Nutrition bulletin, 44(3), 202-215.
CrossRef - Ma, Z., Boye, J. I., & Hu, X. (2018). Nutritional quality and techno-functional changes in raw, germinated and fermented yellow field pea (Pisumsativum L.) upon pasteurization. LWT, 92, 147-154.
CrossRef - Mondor, M. (2020). Pea. Pulses: Processing and Product Development, 245-273.
CrossRef - Melchior, S., Calligaris, S., Bisson, G., &Manzocco, L. (2020). Understanding the impact of moderate-intensity pulsed electric fields (MIPEF) on structural and functional characteristics of pea, rice and gluten concentrates. Food and Bioprocess Technology, 13, 2145-2155.
CrossRef - Ahmed, E. A. A. S., Abo El-Azam, N. A., El-Behairy, U. A. A., & Abdallah, M. M. F. (2018). Organic pea sprout in door to improve pie quality. Arab Universities Journal of Agricultural Sciences, 26(Special issue (2B)), 1449-1460.
CrossRef - Meftahizade, H., Sargsyan, E., &Moradkhani, H. (2010). Investigation of antioxidant capacity of Melissa officinalis L. essential oils. J Med Plant Res, 4(14), 1391-5.
- Cunniff, P., & Washington, D. (1997). Official methods of analysis of AOAC international. J. AOAC Int, 80(6), 127A.
CrossRef - Prosky, L., Asp, N. G., Furda, I., Devries, J. W., Schweizer, T. F., & Harland, B. F. (1984). Determination of total dietary fiber in foods, food products, and total diets: interlaboratory study. Journal of the Association of Official Analytical Chemists, 67(6), 1044-1052.
CrossRef - Livesey, G. (1995). Metabolizable energy of macronutrients. The American journal of clinical nutrition, 62(5), 1135S-1142S.
CrossRef - Lalas, S., &Tsaknis, J. (2002). Characterization of Moringaoleifera seed oil variety “Periyakulam 1”. Journal of food composition and analysis, 15(1), 65-77.
CrossRef - Klein, B. P., & Perry, A. K. (1982). Ascorbic acid and vitamin A activity in selected vegetables from different geographical areas of the United States. Journal of Food Science, 47(3), 941-945.
CrossRef - Rutkowski, M., &Grzegorczyk, K. (2007). Modifications of spectrophotometric methods for antioxidative vitamins determination convenient in analytic practice. ActaScientiarumPolonorum Technologia Alimentaria, 6(3), 17-28.
CrossRef - Park, Y. K., Koo, M. H., Ikegaki, M., &Contado, J. O. S. E. (1997). Comparison of the flavonoid aglycone contents of Apismelliferapropolis from various regions of Brazil. Arq. Biol. Tecnol, 97-106.
- Singleton, V. L., & Rossi, J. A. (1965). Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. American journal of Enology and Viticulture, 16(3), 144-158.
CrossRef - Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M., & Rice-Evans, C. (1999). Antioxidant activity applying an improved ABTS radical cationdecolorization assay. Free radical biology and medicine, 26(9-10), 1231-1237.
CrossRef - Blois, M. S. (1958). Antioxidant determinations by the use of a stable free radical. Nature, 181(4617), 1199-1200.
CrossRef - Matthews, D. R., Hosker, J. P., Rudenski, A. S., Naylor, B. A., Treacher, D. F., & Turner, R. C. (1985). Homeostasis model assessment: insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. diabetologia, 28, 412-419.
CrossRef - Goren, I., Pfeilschifter, J., & Frank, S. (2003). Determination of leptin signaling pathways in human and murine keratinocytes. Biochemical and Biophysical Research Communications, 303(4), 1080-1085.
CrossRef - Exon, J. H. (2006). A review of the toxicology of acrylamide. Journal of Toxicology and Environmental Health, Part B, 9(5), 397-412.
CrossRef - Reeves, P. G., Nielsen, F. H., & Fahey Jr, G. C. (1993). AIN-93 purified diets for laboratory rodents: final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. The Journal of nutrition, 123(11), 1939-1951.
CrossRef - McLaughlan, J. M., & Campbell, J. A. (1964). Methodology of protein evaluation. Mammalian protein metabolism, 3, 391-422.
CrossRef - Hegsted, D. M., Mills, R. C., Elvehjem, C. A. F., & Hart, E. B. (1941). Choline in the nutrition of chicks. Journal of biological chemistry, 138, 459-466.
CrossRef - Chapman, D. G., Castillo, R., & Campbell, J. A. (1959). Evaluation of protein in foods: 1. A method for the determination of protein efficiency ratios. Canadian Journal of Biochemistry and Physiology, 37(5), 679-686.
CrossRef - Schermer, S. (1967). The blood morphology of laboratory animal Longmans. Green and Co. Ltd, 350.
- Trinder, P. (1969): Glucose. Ann. Clin. Biochem., (62):24-33.
CrossRef - Reitman, S., & Frankel, S. (1957). A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. American journal of clinical pathology, 28(1), 56-63.
CrossRef - Henry, R. J. (1974). Clinical Chemist. Principles and Techniqus, 2nd Edition, Hagerstown (MD), Harcer, Row, 882.
CrossRef - Weichselbaum, C. T. (1946). An accurate and rapid method for the determination of proteins in small amounts of blood serum and plasma. American journal of clinical pathology, 16(3_ts), 40-49.
- Tietz, N. W., Rinker, A. D., & Shaw, L. M. (1983). IFCC methods for the measurement of catalytic concentration of enzymes Part 5. IFCC method for alkaline phosphatase (orthophosphoric-monoester phosphohydrolase, alkaline optimum, EC 3.1. 3.1). Journal of clinical chemistry and clinical biochemistry. Zeitschrift fur klinischeChemie und klinischeBiochemie, 21(11), 731-748.
CrossRef - Doumas, B. T., Watson, W. A., & Biggs, H. G. (1971). Albumin standards and the measurement of serum albumin with bromcresol green. Clinicachimicaacta, 31(1), 87-96.
CrossRef - Barham, D., &Trinder, P. (1972). Enzymatic determination of uric acid. Analyst, 97, 142-145.
CrossRef - Patton, C. J., & Crouch, S. R. (1977). Spectrophotometric and kinetics investigation of the Berthelot reaction for the determination of ammonia. Analytical chemistry, 49(3), 464-469.
CrossRef - Allain, C. C., Poon, L. S., Chan, C. S., Richmond, W. F. P. C., & Fu, P. C. (1974). Enzymatic determination of total serum cholesterol. Clinical chemistry, 20(4), 470-475.
CrossRef - Fossati, P., &Prencipe, L. (1982). Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clinical chemistry, 28(10), 2077-2080.
CrossRef - Hamoda, A. M., El-Bedawey, A. A., Khalil, A. H., &Bakry, A. A. (2018). Biological evaluation of balady bread as affected by replacing wheat flour with different levels of date fiber. Menoufia Journal of Food and Dairy Sciences, 3(2), 15-26.
CrossRef - Lopes-Virella, M. F., Stone, P., Ellis, S., & Colwell, J. A. (1977). Cholesterol determination in high-density lipoproteins separated by three different methods. Clinical chemistry, 23(5), 882-884.
CrossRef - Satoh, K. (1978) Determination of total antioxidant capacity (TAC) Clinica. ChimicaActa. Published online:37-90.
- Ohkawa, H., Ohishi, W., &Yagi, KA. (1979) Determination of MDA. Biochem. Published online 1979:95-351.
CrossRef - Sun, Y. I., Oberley, L. W., & Li, Y. (1988). A simple method for clinical assay of superoxide dismutase. Clinical chemistry, 34(3), 497-500.
CrossRef - Koracevic, D.C. (2001) Determination of glutathione s-transferases (GSTs). JCliPathol; 54:356-361.
CrossRef - Sen, C. K. (2001). Antioxidant and redox regulation of cellular signaling: introduction. Medicine and science in sports and exercise, 33(3), 368-370.
CrossRef - Diego, S. (2011). Oxiselect TM Catalase Activity Assay Colorimetric. Cell Biolabs.
- Tabor, H., & Tabor, C. W. (Eds.). (1970). Metabolism of Amino Acids and Amines: Part A. Academic Press.
- Rosano, T. G., Clayson, K. J., &Strandjord, P. E. (1976). Evaluation of adenosine 5′-monophosphate and fluoride as adenylate kinase inhibitors in the creatine kinase assay. Clinical chemistry, 22(7), 1078-1083.
CrossRef - Blaauwen, D. D., Poppe, W. A., &Tritschler, W. (1983). Cholinesterase (EC 3.1. 1.8) mitButyrylthiocholin-iodidalsSubstrat: Referenzwerte in Abhängigkeit von Alter und Geschlechtunter Berücksichtigung hormonaler Einflüsse und Schwangerschaft.
CrossRef - Ratcliffe, M. J. (2016). Encyclopedia of immunobiology. Academic Press.
- Drury, R. A. B., & Wallington, E. A. (1980). Carleton’s histological technique 5th ed. New York: Churchill Livingstone.
- Taiz, L., Zeiger, E., Møller, I. M., & Murphy, A. (2015). Plant physiology and development (No. Ed. 6). Sinauer Associates Incorporated.
- Januškevičius, A., Januškevičienė, G., &Andrulevičiūtė, V. (2012). Chemical composition and energetic values of selected vegetable species in Lithuanian supermarkets. VeterinarijairZootechnika, 58(80).
- Campas-Baypoli, O. N., Sánchez-Machado, D. I., Bueno-Solano, C., Núñez-Gastélum, J. A., Reyes-Moreno, C., &López-Cervantes, J. (2009). Biochemical composition and physicochemical properties of broccoli flours. International Journal of Food Sciences and Nutrition, 60(sup4), 163-173.
CrossRef - Barrón-Yánez, M. R., Villanueva-Verduzco, C., García-Mateos, M. R., & Colinas-León, M. T. (2009). Nutrient value and saponin content of huauzontle (ChenopodiumnuttalliaeSaff.), zucchini (Cucurbita pepo L.), canola (Brassica napus L.) and amaranto (Amaranthusleucocarpus S. Watson syn. hypochondriacus L.) sprouts. RevistaChapingo. Seriehorticultura, 15(3), 237-243.
CrossRef - Mazza, G. (2000). Alimentosfuncionales. Aspectosbioquímicos y de procesados, Ed. Acribia, 93-140.
- Anwar, D.A. (2016): Effect of number of harvests on the production and nutritive value of mushroom fruits, sprout vegetables and their residues. Ph.D. Thesis of Fac. Agric. Ain Shams Univ., Cairo, Egypt, pp. 93-106.
- Santos, J., Herrero, M., Mendiola, J. A., Oliva-Teles, M. T., Ibáñez, E., Delerue-Matos, C., & Oliveira, M. B. P. P. (2014). Assessment of nutritional and metabolic profiles of pea shoots: The new ready-to-eat baby-leaf vegetable. Food Research International, 58, 105-111.
CrossRef - Pérez‐Balibrea, S., Moreno, D. A., &García‐Viguera, C. (2008). Influence of light on health‐promoting phytochemicals of broccoli sprouts. Journal of the Science of Food and Agriculture, 88(5), 904-910.
CrossRef - Herr, I., &Büchler, M. W. (2010). Dietary constituents of broccoli and other cruciferous vegetables: implications for prevention and therapy of cancer. Cancer treatment reviews, 36(5), 377-383.
CrossRef - Ahmed, M.Y. (2014): Effect of germination methods on sprout production and active ingredient contents of some cruciferous crops. M.Sc. Thesis of Fac. Agric. Ain Shams Univ., Cairo, Egypt, pp. 37-51.
- Shah, S. A., Zeb, A., Masood, T., Noreen, N., Abbas, S. J., Samiullah, M., …& Muhammad, A. (2011). Effects of sprouting time on biochemical and nutritional qualities ofMungbean varieties. African Journal of Agricultural Research, 6(22), 5091-5098.
- Severini, C., Giuliani, R., De Filippis, A., Derossi, A., & De Pilli, T. (2016). Influence of different blanching methods on colour, ascorbic acid and phenolics content of broccoli. Journal of food science and technology, 53, 501-510.
CrossRef - Fernandez-Orozco, R., Frias, J., Zielinski, H., Piskula, M. K., Kozlowska, H., & Vidal-Valverde, C. (2008). Kinetic study of the antioxidant compounds and antioxidant capacity during germination of Vignaradiata cv. emmerald, Glycine max cv. jutro and Glycine max cv. merit. Food chemistry, 111(3), 622-630.
CrossRef - Toker, Z. (2009). Variation of total hypericin, phenolic and flavonoid compounds in Hypericumtriquetrifolium during its phenological cycle. Pharmaceutical Biology, 47(4), 285-288.
CrossRef - Pérez-Balibrea, S., Moreno, D. A., &García-Viguera, C. (2011). Genotypic effects on the phytochemical quality of seeds and sprouts from commercial broccoli cultivars. Food Chemistry, 125(2), 348-354.
CrossRef - Paśko, P., Bartoń, H., Zagrodzki, P., Gorinstein, S., Fołta, M., &Zachwieja, Z. (2009). Anthocyanins, total polyphenols and antioxidant activity in amaranth and quinoa seeds and sprouts during their growth. Food chemistry, 115(3), 994-998.
CrossRef - Ying, M., Azlan, A., Al-Sheraji, S. H., Hassan, F. A., & Prasad, K. N. (2013). Antioxidant activities and total phenolic content in germinated and non-germinated legume extracts following alkaline-acid hydrolysis. Pakistan journal of Nutrition, 12(12), 1036.
CrossRef - Bagetti, M., Facco, E. M. P., Piccolo, J., Hirsch, G. E., Rodriguez-Amaya, D., Kobori, C. N., … &Emanuelli, T. (2011). Physicochemical characterization and antioxidant capacity of pitanga fruits (Eugenia uniflora L.). Food Science and Technology, 31, 147-154.
CrossRef - Baenas, N., Moreno, D. A., &García-Viguera, C. (2012). Selecting sprouts of Brassicaceae for optimum phytochemical composition. Journal of agricultural and food chemistry, 60(45), 11409-11420.
CrossRef - Gawlik-Dziki, U., Świeca, M., Dziki, D., &Sugier, D. (2013). Improvement of nutraceutical value of broccoli sprouts by natural elicitors. ActaScientiarumPolonorumHortorumCultus, 12(1), 129-140.
- Cousin, R. (1997). Peas (Pisumsativum L.). Field crops research, 53(1-3), 111-130.
CrossRef - Yener, Y., Kalıpcı, E., Öztaş, H., Aydın, A. D., &Yıldız, H. (2013). Possible neoplastic effects of acrylamide on rat exocrine pancreas. Biotechnic&Histochemistry, 88(1), 47-53.
CrossRef - Huang, M., Zhuang, P., Jiao, J., Wang, J., & Zhang, Y. (2018). Association of acrylamide hemoglobin biomarkers with obesity, abdominal obesity and overweight in general US population: NHANES 2003–2006. Science of the Total Environment, 631, 589-596.
CrossRef - Douny, C., Widart, J., Maghuin-Rogister, G., De Pauw, E., &Scippo, M. L. (2012). Quantification of acrylamide in various Belgian potato products using solid phase extraction and liquid chromatography tandem mass spectrometry detection. Food and Public Health, 2(5).
CrossRef - Wang, B., Qiu, W., Yang, S., Cao, L., Zhu, C., Ma, J., …& Chen, W. (2020). Acrylamide exposure and oxidative DNA damage, lipid peroxidation, and fasting plasma glucose alteration: association and mediation analyses in Chinese urban adults. Diabetes Care, 43(7), 1479-1486.
CrossRef - Zhang, P., Pan, H., Wang, J., Liu, X., & Hu, X. (2014). Telomerase activity-independent function of telomerase reverse transcriptase is involved in acrylamide-induced neuron damage. Biotechnic&Histochemistry, 89(5), 327-335.
CrossRef - Adam, T. C., &Epel, E. S. (2007). Stress, eating and the reward system. Physiology & behavior, 91(4), 449-458.
CrossRef - Marroqui, L., Gonzalez, A., Neco, P., Caballero-Garrido, E., Vieira, E., Ripoll, C., …& Quesada, I. (2012). Role of leptin in the pancreatic β-cell: effects and signaling pathways. Journal of molecular endocrinology, 49(1), R9-R17.
CrossRef - Hemphill, D. (2011): Commercial vegetable production guides: Peas, Western Oregon. Oregon State University.
- Teng, W., Li, Y., Du, M., Lei, X., Xie, S., & Ren, F. (2019). Sulforaphane prevents hepatic insulin resistance by blocking serine palmitoyltransferase 3-mediated ceramide biosynthesis. Nutrients, 11(5), 1185.
CrossRef - Lala, V., Goyal, A., Bansal, P., & Minter, D. A. (2020). Liver Function Tests. Treasure Island, FL.
- Guerrero-Beltrán, C. E., Calderón-Oliver, M., Pedraza-Chaverri, J., &Chirino, Y. I. (2012). Protective effect of sulforaphane against oxidative stress: recent advances. Experimental and Toxicologic Pathology, 64(5), 503-508.
CrossRef - Dueñas, M., Estrella, I., & Hernández, T. (2004). Occurrence of phenolic compounds in the seed coat and the cotyledon of peas (Pisumsativum L.). European Food Research and Technology, 219, 116-123.
CrossRef - BelhadjBenziane, A., DilmiBouras, A., Mezaini, A., Belhadri, A., &Benali, M. (2019). Effect of oral exposure to acrylamide on biochemical and hematologic parameters in Wistar rats. Drug and chemical toxicology, 42(2), 157-166.
CrossRef - Jabbar, S. A. (2011). Study the effect of acrylamide on some physiological and histological properties of rabbits kidney who drenched with Flavonoids that extracted from grape seeds. journal of kerbala university, 9(2).
- Eggler, A. L., &Savinov, S. N. (2013). Chemical and biological mechanisms of phytochemical activation of Nrf2 and importance in disease prevention. 50 Years of Phytochemistry Research: Volume 43, 121-155.
CrossRef - Rawi, S. M., Marie, M. A. S., Fahmy, S. R., & El-Abied, S. A. (2012). Hazardous effects of acrylamide on immature male and female rats. Afr J Pharm Pharmacol, 6(18), 1367-86.
CrossRef - Al-Agele, F. A. L., &Khudiar, K. K. (2016). Effect of acrylamide and fructose on some parameters related to metabolic syndrome in adult male rats. Iraqi J. Vet. Med, 40(1), 125-135.
CrossRef - Mahmood, S. A. F., Amin, K., Rahman, H. S., & Othman, H. H. (2016). The pathophysiological effects of acrylamide in Albino Wister rats. International Journal of Medical Research & Health Sciences, 5(7), 42-48.
- Armah, C. N., Derdemezis, C., Traka, M. H., Dainty, J. R., Doleman, J. F., Saha, S., …&Mithen, R. F. (2015). Diet rich in high glucoraphanin broccoli reduces plasma LDL cholesterol: Evidence from randomised controlled trials. Molecular nutrition & food research, 59(5), 918-926.
CrossRef - Sudini, K., Diette, G. B., Breysse, P. N., McCormack, M. C., Bull, D., Biswal, S., … & Matsui, E. C. (2016). A randomized controlled trial of the effect of broccoli sprouts on antioxidant gene expression and airway inflammation in asthmatics. The Journal of Allergy and Clinical Immunology: In Practice, 4(5), 932-940.
CrossRef - Yeap, S. K., Beh, B. K., Ho, W. Y., MohdYusof, H., Mohamad, N. E., Ali, N. M., … & Long, K. (2015). In vivo antioxidant and hypolipidemic effects of fermented mung bean on hypercholesterolemic mice. Evidence-Based Complementary and Alternative Medicine, 2015.
CrossRef - Liu, D., Guan, X., Huang, K., Li, S., Liu, J., Yu, W., &Duan, R. (2019). Protective effects of mung bean (Vignaradiata L.) and pea (Pisumsativum L.) against high‐fat‐induced oxidative stress. Food Science & Nutrition, 7(12), 4063-4075.
CrossRef - Pradeep, K., Mohan, C. V. R., Gobianand, K., &Karthikeyan, S. (2007). Silymarin modulates the oxidant–antioxidant imbalance during diethylnitrosamine induced oxidative stress in rats. European journal of pharmacology, 560(2-3), 110-116.
CrossRef - Kohen, R., &Nyska, A. (2002). Invited review: oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. Toxicologic pathology, 30(6), 620-650.
CrossRef - Shapiro, T. A., Fahey, J. W., Wade, K. L., Stephenson, K. K., &Talalay, P. (2001). Chemoprotectiveglucosinolates and isothiocyanates of broccoli sprouts: metabolism and excretion in humans. Cancer Epidemiology Biomarkers & Prevention, 10(5), 501-508.
- Ge, J., Sun, C. X., Corke, H., Gul, K., Gan, R. Y., & Fang, Y. (2020). The health benefits, functional properties, modifications, and applications of pea (Pisumsativum L.) protein: Current status, challenges, and perspectives. Comprehensive Reviews in Food Science and Food Safety, 19(4), 1835-1876.
CrossRef - Mejri, F., Khoud, H. B., Njim, L., Baati, T., Selmi, S., Martins, A., …& Hosni, K. (2019). In vitro and in vivo biological properties of pea pods (Pisumsativum L.). Food Bioscience, 32, 100482.
CrossRef - Devi, J., Sanwal, S. K., Koley, T. K., Mishra, G. P., Karmakar, P., Singh, P. M., & Singh, B. (2019). Variations in the total phenolics and antioxidant activities among garden pea (Pisumsativum L.) genotypes differing for maturity duration, seed and flower traits and their association with the yield. ScientiaHorticulturae, 244, 141-150.
CrossRef - Zhao, T., Su, W., Qin, Y., Wang, L., & Kang, Y. (2020). Phenotypic diversity of pea (Pisumsativum L.) varieties and the polyphenols, flavonoids, and antioxidant activity of their seeds. Ciência Rural, 50, e20190196.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








