
Introduction
Fruits are perishable by nature and their low pH values give fungi a competitive advantage over most bacteria and yeasts, thereby making them more prone to fungal onslaught. Among the most encountered species of fungi responsible for the spoilage of fruits and derived products, belong to the genus Penicillium1.
A novel emerging fungal pathogen often seen on moldy citrus fruits, which hails from the above stated genus is Penicillium citrinum1,2. It has been isolated from cucumber, dried wine fruits, grapes, lemon, oranges, persian lime, sweet lime and tomatoes2-6. Occurrence of P. citrinum has also been reported on capsicum sp. during pre-harvest, harvest, and post- harvest stages7, where it serves as the predominant source of mycotoxin citrinin, which poses serious health concerns and economic threat, globally, due to its potential hepatotoxicity and nephrotoxicity to humans and adverse impact on valuable agricultural commodities8.
Another broad spectrum fungal pathogen is Penicillium expansum, and it is the only Penicillium species that can express pathogenicity over a variety of fruits, viz., apricots, avocadoes, citrus, grapes, kiwi fruits, mangoes, nectarines, plums, pome fruit, stone fruit, strawberries and tomatoes9,10. P. expansum is also well known for causing the rotting of apples and pears, and is mainly responsible for the production of mycotoxin patulin in juices extracted from the rotted fruits11,12. Consumption of patulin contaminated food products may lead to various health hazards in humans involving gastrointestinal disorders, nausea and pulmonary congestion along with possible carcinogenic, genotoxic, immunotoxic, neurotoxic and teratogenic effects13.
Till date, use of synthetic chemicals is prevalent to control the growth of microbes for the preservation of raw agricultural commodities and processed food products, which is continuously being challenged by consumers and scientific community due to their possible toxicity, harmful effect on environment, high cost and development of resistant races of pathogens14. Hence, there is an increasing demand of natural preservatives, those are safe, cost effective and eco- friendly15,16. Spices being of plant origin, undoubtedly, lie under this category.
India is the world’s largest consumer, exporter, and producer of spices17. Despite the fact that use of spices for the preservation of food products dates back history, these precious horticultural crops are widely being rediscovered for their antioxidant and antimicrobial properties, for last few decades. Although, various findings have been reported on the antimicrobial activities of spice extracts and some spice phtyochemicals, such as allicin, cinnamic aldehyde and eugenol etc., towards several pathogenic microbes of animals and humans (Aspergillus spp., Bacillus spp., Enterococcus faecalis, Escherichia coli, Listeria monocytogenes, Pseudomonas spp., Salmonella spp., Shigella sonnei, Staphylococcus aureus, Vibrio parahaemolyticus and many others)18-23. It has also been well documented that EOs and their components bind with the lipid bilayer of the bacterial cell membrane and mitochondria, and alter the cell structures23. But, there is scanty information available on the inhibitory effect of spices on fungi causing the spoilage of fruits and vegetables.
Considering above aspects in mind, present study was designed to investigate growth inhibitory potentials of AEs, EOs and PD forms of some Indian culinary spices, towards phytopathogenic Penicillium citrinum (MTCC2553) and Penicillium expansum (MTCC2006). In comparison to previous researches, which mainly focused on antimicrobial properties of spice EOs, we screened PD spices as well, because spices in their later form are most commonly used in day-to-day domestic culinary practices in India as an important seasoning ingredient. Secondly, PD spices cost less as compared to their corresponding EOs, those involve expensive and tedious methods of production.
Therefore, this study would serve as harbinger to establish spices as ‘green antimicrobials’ to extend the shelf life of fruits and vegetables and derived products, which would be contributing to agricultural sector and food processing industry, equally.
Materials and Methods
Procurement and preparation of spice samples
PD spice samples
Plant parts of Allium sativum (As), Mentha piperita (Mp) and Zingiber officinale (Zo), were procured in their fresh forms, from a local grocery shop. Ocimum sanctum (Os) leaves were hand plucked from a home grown plant. Peels of As bulbs and Zo rhizomes were removed manually with knife. Fresh forms of aforementioned 4 spices were washed with distilled water to remove the extraneous matter, and were subsequently dried under ambient conditions in shade.
Dried plant parts of Brassica juncea (Bj), Cinnamomum cassia (Blume) (Ccb), Cuminum cyminum (Ccm), Curcuma longa (Cl), Syzygium aromaticum (Sa) and Trigonella foenum-graecum (Tf-g) were purchased from a spice wholesaler of local market, and were manually cleaned to remove extraneous material.
Dried forms of spices thus obtained, were ground to powdered (PD) forms with the help of a grinder in the laboratory and were stored in hermatically sealed jars till their further use. Botanical features of Indian culinary spices used in current study are presented in Table 1.
EOs of spices
EOs of spices were purchased from Aroma Chemicals Pvt. Limited, India. Company assured the purity of the spice EOs to be more than 98.0 %.
AEs of spices
Spice AEs were prepared by the method described elsewhere21. All the AEs were collected in sterilized glass vials, and were further used within 2 h. of their preparation, at various concentration levels as per the requirement of experiment.
Table 1: Botanical features of Indian culinary spices used in current study
|
Botanical names |
Indian names |
English names |
Family |
Plant parts of spices used |
|
Allium sativum L. |
Lehasun |
Garlic |
Liliaceae |
Bulbs |
|
Brassica juncea L. |
Sarson |
Brown Mustard |
Crucifereae |
Seeds |
|
Cinnamomum cassia (Blume) |
Dalchini |
Cinnamon |
Lauraceae |
Bark |
|
Cuminum cyminum L. |
Jeera |
Cumin |
Apiaceae |
Seeds |
|
Curcuma longa L. |
Haldi |
Turmeric |
Zingiberaceae |
Rhizomes |
|
Mentha piperita L. |
Paudina |
Mint |
Labiateae |
Leaves |
|
Ocimum sanctum L. |
Tulsi |
Holy Basil |
Lamiaceae |
Leaves |
|
Syzygium aromaticum L. |
Laung |
Clove |
Myrtaceae |
Flower Buds |
|
Trigonella foenum-graecum L. |
Methi |
Fenugreek |
Leguminosae |
Seeds |
|
Zingiber officinale L. |
Adarak |
Ginger |
Zingiberaceae |
Rhizomes |
Fungal strains and growth conditions
Pure cultures of Penicillium citrinum (MTCC 2553) and Penicillium expansum (MTCC 2006) were procured from Microbial Type Culture Collection (MTCC), Chandigarh, India Potato dextrose broth medium (PDB) and Potato dextrose agar medium (PDA) (Hi Media, India) were used for the cultivation of both the fungi (temperature of incubation : 25o C ; duration of incubation : 48 h-96 h), as per the recommendations of MTCC. Initially, the growth of both the fungi appeared as white mycelium on PDA that turned to greenish to bluish green in colour as the spores matured.
Preparation of fungal inoculum
For the preparation of inoculum, bluish green spores of P. citrinum and P. expansum were harvested from 15 days old slants by adding 0.05% of Tween 80 ((Central Drug House (CDH), India)). These harvested spores were quantified by a haemocytometer to adjust at 107 spores/mL.
Screening inhibitory activities of spice samples
Spice agar method
Spice agar method24 was used for the determination of inhibitory potentials of PD spice samples. Freshly prepared fungal inoculum (100 µL) was evenly spread over the surface of solidified PDA petri plates previously supplemented with various concentrations of PD spices (0.1 – 5.0 (%,w/v)). Seeded petri plates were incubated at 25oC for 30 days (total incubation period), and were examined for the growth of fungi at a regular interval of 12 h., consistently. The time for the initiation of fungal growth on petri plates were recorded. A control set of experiment (without any PD spice sample), was also conducted, parallelly.
From the results of spice agar method, minimum inhibitory concentrations (MICs) of PD spice samples were derived. For the sake, concentrations of spices (COS (%, w/v)) were plotted on the x-axis and days of inhibition (DOI) were plotted on the y-axis of graph (Figure 1). Then, 40%, 60% and 80% levels of total incubation period (30 days), were calculated. Afterwards, from each referred level, a horizontal line was drawn to intersect the curve. Subsequently, a perpendicular line was drawn from the point of intersection, which corresponded to the concentration of spice sample. MIC was the concentration of PD spice sample which did not allow the fungi to grow up to 80% level, i.e., 24 days.
Impregnated paper disc method
Impregnated paper disc method, narrated by someone else25 was opted for screening antifungal activities of AEs and EOs of spices. Results were presented as net zones of inhibition (mm) after subtracting diameter of filter paper discs (6mm) from the diameter of measured inhibitory zones.
Broth dilution assay
Broth dilution assay25, was used to determine minimum inhibitory concentrations (MICs) of AEs and EOs (which exhibited distinct inhibitory zones during impregnated disc method). PDB containing 2000 µL/mL of test EOs were serially diluted twofold each with PDB to get concentration levels of 1000, 500, 250, 125, 62.50, 31.25, 15.62, 7.81, 3.90, 1.95, 0.97, 0.48, 0.24, 0.12, 0.06 µL/mL. Then, 100 µL of inoculum of test fungi was added to them. Mixtures thus obtained were incubated at 25ºC for 96 h. After the stipulated incubation period, 100 µL of above mixture was spread on the surface of solidified PDA plates. The later were incubated for 96 h, at 25 ºC to notice the lowest dilution of test samples, which did not show any visible growth of fungi.
*During impregnated paper disc method and broth dilution assay, dimethylsulphoxide (DMSO) (CDH, India), was used as a negative control agent. All the above experiments were conducted thrice, simultaneously.
Statistical analysis
Results of zone inhibition assay were statistically analyzed by using SPSS version 7.5 and were expressed as the mean ± standard deviation.
Results
Inhibitory activities of PD spices
Data presented in Table 2 indicate that PDA petri dishes incorporated with PD forms of Ccb, Ccm and Sa, at their different concentration levels (from 0.1 to 5.0 (%,w/v)), potentially inhibited P. citrinum (Pcit) and P. expansum (Pexp). On the contrary, Bj, Cl, Mp, Os, Tf-g and Zo, up to their highest concentration level of 5.0% (w/v), were found ineffective in inhibiting test fungal strains, and their growth in culture media in the form of white mycelium was observed on the 2nd day of incubation as in control set of petri dishes without any PD spice sample. However, As displayed very weak inhibitory effect only towards Pcit, and former at 5.0% level, delayed the growth of later by 4 days. It is obvious from the data of spice agar method that both the fungi responded in their own peculiar manner towards various concentrations of Ccb, Ccm and Sa. Ccb and Sa at a concentration level as low as 0.1% (w/v), exerted their inhibitory effects towards Pcit, while growth of Pexp was delayed by both the aforestated spices at their 0.2% levels (w/v).
It was also noticed that number of days of inhibition increased with the increase in the concentration of PD spices. And, Sa at concentration levels of 2.0% (w/v) and 3.0% (w/v) delayed the growth of Pcit and Pexp respectively, by 30 days. Similarly, Ccb at 3.0% (w/v) level, hindered the visible growth of both the test fungi throughout the incubation period. On the other hand, Ccm up to 1.5% (w/v) was found to be ineffective towards test fungi, and showed its inhibitory effect at a concentration level of 2.0% (w/v) by impeding the growth of Pcit and Pexp for 5 days and 4 days, respectively. At 5.0% (w/v) level, Ccm delayed the visible growth of Pcit up to 21 days, and that of Pexp up to 17 days , and was thus unable to produce the desired growth inhibitory effect even at its highest stated concentration level.
It is worth mentioning here that different levels of inhibition were generated by different concentrations of referred spices towards both the microbes under question (Figure 1, Table 3). Ccb and Sa produced 40%, 60% and 80% levels of inhibition towards both the fungal strains, while in case of Ccm, 80% level of inhibition was not detected. Though, a positive and direct relation was observed between different levels of inhibition generated and the concentration of PD spices used (Figure 1). The concentration of spice which produced 80% level of inhibition against test fungi was considered as minimum inhibitory concentration (MIC) (Figure 1, Table 3). It is quite noteworthy that lower concentrations of PD spices were required to produce any given level of inhibition against Pcit as compared to Pexp.
On the basis of days of inhibition produced and MIC values obtained, PD spice forms can be put in the following order in terms of their decreasing inhibitory effect towards fungal strains : Pcit: Sa> Ccb> Ccm ; Pexp: Ccb> Sa> Ccm.
Table 2: Effect of different concentrations of PD spices on growth of fungi
|
Spice Conc. (%,w/v) |
Days of Inhibition |
|||||||
|
As |
Ccb |
Ccm |
Sa |
|||||
|
Pcit |
Pexp |
Pcit |
Pexp |
Pcit |
Pexp |
Pcit |
Pexp |
|
|
0.0 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
|
0.1 |
2 |
2 |
3 |
2 |
2 |
2 |
4 |
2 |
|
0.2 |
2 |
2 |
5 |
3 |
2 |
2 |
7 |
3 |
|
0.4 |
2 |
2 |
7 |
4 |
2 |
2 |
9 |
4 |
|
0.6 |
2 |
2 |
8 |
7 |
2 |
2 |
11 |
6 |
|
0.8 |
2 |
2 |
12 |
9 |
2 |
2 |
16 |
9 |
|
1.0 |
2 |
2 |
14 |
12 |
2 |
2 |
20 |
11 |
|
1.5 |
2 |
2 |
18 |
17 |
2 |
2 |
28 |
14 |
|
2.0 |
2 |
2 |
23 |
21 |
5 |
4 |
>30 |
21 |
|
2.5 |
2 |
2 |
29 |
27 |
8 |
5 |
>30 |
26 |
|
3.0 |
2 |
2 |
>30 |
>30 |
10 |
8 |
>30 |
>30 |
|
3.5 |
2 |
2 |
>30 |
>30 |
14 |
11 |
>30 |
>30 |
|
4.0 |
2 |
2 |
>30 |
>30 |
16 |
12 |
>30 |
>30 |
|
4.5 |
4 |
2 |
>30 |
>30 |
19 |
15 |
>30 |
>30 |
|
5.0 |
5 |
2 |
>30 |
>30 |
21 |
17 |
>30 |
>30 |
As: Allium sativum, Ccb: Cinnamomum cassia (Blume), Ccm: Cuminum cyminum Sa: Syzygium aromaticum, Pcit: Penicillium citrinum, Pexp: Penicillium expansum
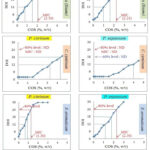 |
Figure 1: Levels of inhibition generated by PD spices |
Table 3 : Levels of inhibition generated by PD spices
|
Levels of Inhibiton |
Spice concentration (%, w/v) |
|||||
|
Ccb |
Ccm |
Sa |
||||
|
Pcit |
Pexp |
Pcit |
Pexp |
Pcit |
Pexp |
|
|
40% |
0.80 |
1.00 |
3.28 |
4.00 |
0.70 |
1.20 |
|
60% |
1.50 |
1.60 |
4.30 |
NOB |
0.90 |
1.60 |
|
80% |
2.10 |
2.23 |
NOB |
NOB |
1.30 |
2.30 |
Ccb: Cinnamomum cassia (Blume), Ccm: Cuminum cyminum, Sa: Syzygium aromaticum, Pcit: Penicillium citrinum, Pexp: Penicillium expansum, NOB: Not observed
Inhibitory activities of AEs and EOs of spices
Results (Table 4) of impregnated paper disc method show that petriplates poured with AEs of test spices did not show any growth inhibitory zone towards Pcit and Pexp. Same response of test fungi was observed towards EOs of Cl, Tf-g and Zo, and also with the control set of petriplates incorporated with DMSO. Contrastingly, EOs of As, Bj, Ccb, Ccm, Os, Mp and Sa displayed distinct zones of inhibition (mm) of varying diameters towards both the fungi under observation. It is obvious from the data that Pcit gave wider growth inhibitory zones as compared to Pexp towards all the spice EOs. Based on the measured diameters of inhibitory zones, the following ranking of spice EOs in their descending order can be made towards fungal strains :
Pcit : Ccm> Sa> Ccb> Bj> As> Mp> Os ; Pexp : Sa> Bj> As= Ccm> Ccb> Mp> Os.
MICs of EOs (which exhibited inhibitory zones during impregnated paper disc method), were determined using broth dilution assay, and are enlisted in Table 5. Data revealed that MIC values of EOs towards Pcit ranged from 3.90 µL/mL- 62.50 µL/mL, and towards Pexp, MIC values varied from 7.81 µL/mL- 62.50 µL/mL. On the basis of MIC values obtained, effectivity of spice EOs in descending order towards fungi can be put as :
Pcit : Ccb= Ccm= Sa> As= Bj= Mp> Os ; Pexp : Ccb> As= Bj= Ccm= Sa> Mp> Os.
Table 4: Inhibitory activities of AEs and EOs
|
Spices |
Zones of Inhibition (mm) |
|||||
|
DMSO (5µL) |
EO (5µL) |
AE (10µL) |
||||
|
Pcit |
Pexp |
Pcit |
Pexp |
Pcit |
Pexp |
|
|
As |
NOB |
NOB |
27.10±0.56 |
24.00±0.21 |
NOB |
NOB |
|
Bj |
NOB |
NOB |
40.50±0.09 |
32.00±0.12 |
NOB |
NOB |
|
Ccb |
NOB |
NOB |
41.50±0.16 |
19.50±0.29 |
NOB |
NOB |
|
Ccm |
NOB |
NOB |
42.10±0.51 |
24.10±1.00 |
NOB |
NOB |
|
Cl |
NOB |
NOB |
NOB |
NOB |
NOB |
NOB |
|
Mp |
NOB |
NOB |
12.50±0.33 |
12.00±0.38 |
NOB |
NOB |
|
Os |
NOB |
NOB |
10.30±0.57 |
10.10±0.12 |
NOB |
NOB |
|
Sa |
NOB |
NOB |
42.00±0.67 |
34.00±0.50 |
NOB |
NOB |
|
Tf-g |
NOB |
NOB |
NOB |
NOB |
NOB |
NOB |
|
Zo |
NOB |
NOB |
NOB |
NOB |
NOB |
NOB |
As: Allium sativum, Bj: Brassica juncea, Ccb: Cinnamomum cassia (Blume),
Ccm: Cuminum cyminum, Cl: Curcuma longa, Mp: Mentha piperita,
Os: Ocimum sanctum, Sa: Syzygium aromaticum, Tf: Trigonella foenum-graecum, Zo: Zingiber officinale, Pcit: Penicillium citrinum, Pexp: Penicillium expansum, NOB: Not Observed, DMSO: Dimethylsulphoxide.
Table 5: Minimum inhibitory concentrations (MICs) of EOs
|
Spices |
MICs |
|||
|
DMSO |
EOs |
|||
|
Pcit |
Pexp |
Pcit |
Pexp |
|
|
As |
NOB |
NOB |
7.81 |
15.62 |
|
Bj |
NOB |
NOB |
7.81 |
15.62 |
|
Ccb |
NOB |
NOB |
3.90 |
7.81 |
|
Ccm |
NOB |
NOB |
3.90 |
15.62 |
|
Mp |
NOB |
NOB |
15.62 |
62.50 |
|
Os |
NOB |
NOB |
31.25 |
62.50 |
|
Sa |
NOB |
NOB |
3.90 |
15.62 |
As: Allium sativum, Bj: Brassica juncea, Ccb: Cinnamomum cassia (Blume),
Ccm: Cuminum cyminum, Mp: Mentha piperita, Os: Ocimum sanctum,
Sa: Syzygium aromaticum, Pcit: Penicillium citrinum, Pexp: Penicillium expansum, NOB: Not Observed, DMSO: Dimethylsulphoxide.
Discussion
Spices are well known for their flavouring and therapeutic properties. Mainstream scientific community readily accepts spices as antimicrobial agents. Present in vitro trials clearly suggest that spice forms under investigation showed marked variability in inhibiting phytopathogenic Pcit and Pexp. As per our observations, EOs of spices arrested the fungal growth most potentially, followed by PD spices, whereas AEs were found totally ineffective. Our findings are in consistency with the results of Hetta et al., those elucidated that EOs and PD forms of some spice plants (black cumin, cinnamon, clove, cumin and marjoram), have antibacterial activities against Bacillus spp. isolated from raw and processed meat, to which they further added that inhibitory potential of EOs was stronger than the PD forms of the same plant26.
Furthermore, results of our study revealed that amongst PD forms of spices, Ccb and Sa, arrested the growth of Pcit and Pexp, most significantly followed by Ccm and As, while Bj, Cl, Mp, Os, Tf-g and Zo, proved ineffectual. This variation in the antifungal activities shown by PD spices may be attributed to their volatile EOs. Rio et al. and many others have postulated that functional properties of spices reside in their volatile EOs, which in turn consist of a myriad of phytochemicals, viz., alcohols (farnesol, menthol), aldehydes (cinnamic aldehyde, cuminic aldehyde), polyphenols (eugenol, resveratrol), terpenoids (thymol) and thiols (allicin and allylisothiocyanate), preferably known as bioactive components27,28,29. Aforestated components are basically responsible for the antimicrobial activities of spices, either by causing degradation of cell wall/ cytoplasmic membrane, leakage of cellular components or by affecting their genetic material, protein synthesis, electron transport system, nutrient uptake, enzymatic activities and energy production inside the cell, adversely29,30,31.
Therefore, low effectivity of As and non effectivity of Bj, Cl, Mp, Os, Tf-g and Zo, in their PD forms in current study, may be either due to their less amount of EOs or due to the substantial loss of volatile antimicrobial components during the process of drying and grinding. It is quite evident from our results, wherein EOs of Bj, Mp and Os exhibited growth inhibitory effects, while PD forms of referred spices did not show any antifungal activity.
Moreover, greater antifungal potential of Sa, Ccb and Ccm may be attributed to their high amount of EOs and chemical composition of EOs. Past researches have demonstrated that Sa comprises 15-20% EO, which is primarily eugenol (80-85%)32. Likewise, Ccb also consists of considerable amount of EO that ranges from 0.9-4.0% with major constituent being cinnamic aldehyde33 , and Ccm contains 2.5-4.5% of EO, and is mainly composed of cuminic aldehyde34. Though, it deserves special attention that amount of EO in Ccb is far less as compared to Sa, but antifungal activities shown by PD forms of these 2 spices towards Pcit and Pexp, were close to each other as per our observations. Similarly, Ccm has more amount of EO as compared to Ccb, but in terms of generation of days of inhibition, PD form of former proved less effective than the later. This discrepancy in results, thus can be attributed to different chemical structures of major bioactive components of EOs of spices and their specific mode of action. Other related factors responsible for greater antifungal potentials of PDCcb and PDSa may include the pH, volatility, molecular weight and diffusion of antimicrobial components in growth medium along with type of microorganism implicated in the study. However, exact mechanisms of action of these bioactive components towards Pcit and Pexp at molecular levels are not yet understood, and would remain a line of future research.
Similar reasons can be given for differential antifungal activities shown by EOs of spices towards Pcit and Pexp, during broth dilution assay and impregnated paper disc method, where EOs of Mp and Os were less potent in inhibiting test fungi as compared to EOs of As, Bj,Ccb, Ccm and Sa, towards both the test fungi. Our results were in agreement with the findings of De-Montijo-Prieto et al., wherein they reported antimicrobial activities of 13 EOs from different herbs, spices, fruits and vegetables towards some common food borne pathogens18.
The altogether inert nature of 10 AEs in current trials could be due to the insolubility of hydrophobic antimicrobial components of EOs in water. It is suggested that the hydrophobicity of the EOs and their bioactive components is the detrimental factor which makes them to get accumulated in the lipid bilayer of the cell membrane and mitochondria, rendering them more permeable to external agents Gonelimali et al. also found that aqueous extracts of Hibiscus sabdariffa and Syzygium aromaticum did not show antimicrobial activity against Candida albicans19.
It was also notable that inhibitory effect of EOs and PD forms was more pronounced against Pcit as compared to Pexp. Though, reasons for greater susceptibility of Pcit need further elucidation.
Conclusion
Concludingly, Ccb, Ccm and Sa, in their two test forms, i.e., EOs and PD forms, inhibited Pcit and remarkably, and hence, may be considered as ‘potent inhibitors’. Conversely, Cl, Tf-g and Zo did not show any growth inhibitory activity , in their any test form, and may be categorized as ‘non inhibitors’. Hence, PD forms of Ccb, Ccm and Sa, being cheap and safe, may find a place as ‘natural antimicrobials’ along with EOs of As, Bj, Ccb, Ccm, Mp, Os and Sa, to retard the growth of Pcit and Pexp in order to extend the shelf life of fruits and processed food products. This study would help to reduce post harvest loss of valuable agricultural commodities and meet the ever increasing consumers’ demand of wholesome food which is natural and free from harmful chemical additives.
Acknowledgement
Authors would like to acknowledge the kind assistance of Professor Neeraj Dilbaghi, chairperson, Department of Bio & Nano Technology, Guru Jambheshwar University of Science and Technology, Hisar-125001, Haryana, India, during the work.
Conflict of Interest
Authors have no conflict of interest in this study.
Funding Sources
This study was the part of research project fuded by University Grants Commission (UGC), New Delhi, India.
References
- Shaikh N.U., Chavan A.M. Isolation and characterization of penicillium sps. from citrus. Int J cur res. 2017; 9: 53465-53466.
- Nosheen A., Anjum T., Jabeen R. Isolation and identification of storage fungi from citrus sampled from major growing areas of Punjab, Pakistan. Int J Agr Biol. 2013; 15: 1283–1288.
- Saleh I., Al-Thani R. Fungal food spoilage of supermarkets’ displayed fruits. Vet World. 2019; 12: 1877-1883. DOI:10.14202/vetworld.2019. 1877-1883
CrossRef - Kim W.K., Sang H.K., Woo S.K., Park M.S., Paul N.C., Hun Yu S. Six species of Penicillium associated with blue mold of grape. Mycobiol. 2007; 35: 180-185. DOI:10.4489/MYCO.2007.35.4.180
CrossRef - Coutinho T.C., Ferreira M.C., Rosa L.H., de Oliveira A.M., de Oliveira E.N. Júnior. Penicillium citrinum and Penicillium mallochii: New phytopathogens of orange fruit and their control using chitosan. Carb Poly. 2020; 234: 01-10. DOI: 10.1016/j.carbpol.2020.115918
CrossRef - Estrada R.R.G., de Jesus Ascencio Valle F., Sanchez J.A.R., Santoy M.C. Use of a marine yeast as a biocontrol agent of the novel pathogen Penicillium citrinum on Persian lime. Emi J Food Agr. 2017; 29: 114-122. DOI: 10.9755/ejfa.2016-09-1273
CrossRef - Frimpong G.K., Adekunle A.A., Ogundipe O.T., Solanki M.K., Sadhasivam S., Sionov E. Identification and toxigenic potential of fungi isolated from Capsicum Peppers. Microorganisms. 2019; 7 : 01-10. DOI: 10.3390/microorganisms7090303
CrossRef - Kamle M., Mahato D.K., Gupta A., Pandhi S., Sharma N., Sharma B. Citrinin mycotoxin contamination in food and feed : Impact on agriculture, human health, and detection and management strategies. Toxins. 2022; 14: 01-25. DOI: 10.3390/toxins14020085
CrossRef - Batta Y.A. Quantitative postharvest contamination and transmission of Penicillium expansum (Link) conidia to nectarine and pear fruit by Drosophila maelanogaster (Meig.) adults. Postharv Bio Technol. 2006; 40: 190-196. DOI:10.1016/j.postharvbio.2006.01.006.
CrossRef - Molinu M.G., Pani G., Venditti T., Dore A., Ladu G., D’Hallewin G. Alternative methods to control postharvest decay caused by Penicillium expansum in plants (Prunus domestica L.). Commun Agr App Biol Sci. 2012; 77: 509-514.
- Ivana V., Duduk N., Vasic M., Nikolic M. Identification of Penicillium expansum causing postharvest blue mold decay of apple fruit. Pestic Phytomed. 2014; 29: 257–266. DOI: 10.2298/PIF1404257V
CrossRef - Kim J., Lee W., Cheong S., Choi J., Ryu J., Choi Y. Identification and characteristics of Penicillium spp. isolated from postharvest decay of pear. Res Plant Dis. 2002; 8: 107-112.
CrossRef - Saleh I., Goktepe I. Health risk assessment of Patulin intake through apples and apple-based foods sold in Qatar. Helion. 2019; 5: 01-07. DOI: 10.1016/j.heliyon2019.e02754
CrossRef - Jimenez M., Maldonado M.I., Rodriguez E.M., Hernandez-Ramirez A., Saggioro E., Carra I., et al. Supported TiO2 solar photocatalysis at semi-pilot scale : degradation of pesticides found in citrus processing industry waste water, reactivity and influence of photogenerated species. J Chem Tech Biotech. 2015; 90: 149–157. DOI: 10.1002/jctb.4299
CrossRef - Talibi I., Boubaker H., Boudyach E., Ben Aoumar A. Alternative methods for the control of postharvest citrus diseases. J App Micro. 2014; 117: 01–17. DOI: 10.1111/jam.12495
CrossRef - Fischer I., Lourenco S., Amorim L. Postharvest diseases in citrus and characterization of the fungal population in Sao Paulo’s wholesale market. Trop Plant Path. 2008; 33: 219–226.
CrossRef - Thomas L., Sanil P.C. Competitiveness in spice export trade from India : A review. J Spices Arom Crops. 2019; 28: 01-19. DOI:10.25081/josac.2019.v28.i1.5738
CrossRef - De-Montijo-Prieto S., Razola-Díaz M.C., Gómez-Caravaca A. M., Guerra-Hernandez EJ, Jiménez-Valera M, Garcia-Villanova B, et al. Essential oils from fruit and vegetables, aromatic herbs, and spices: composition, antioxidant, and antimicrobial activities. Biology. 2021; 10: 01-21. DOI: 10.3390/biology10111091
CrossRef - Gonelimali F.D., Lin J., Miao W., Xuan J., Charles F., Chen M. and Hatab S.R. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front Microb.2018; 9: 01-09. DOI : 10.3389/fmicb.2018.01639
CrossRef - Bhatwalkar S.B., Mondal R., Krishna S.B.N., Adam J.K., Govender P., Anupam R. Antibacterial properties of organosulfur compounds of Garlic (Allium sativum). Front Microb.2021; 12: 01-20. DOI: 10.3389/fmicb.2021.613077
CrossRef - Vasconcelos N.G., Croda J., Simionatto S. Antibacterial mechanisms of cinnamon and its constituents: A review. Microb. Path. 2018; 120: 198-203. DOI:10.1016/micpath2018.04.036
CrossRef - Jayapal V. Antimicrobial activity of eugenol against human pathogenic bacteria by minimal inhibitory concentration, minimal bactericidal concentration and disc-diffusion methods. Int J Pharm Sci Res. 2021; 12: 330-335. DOI: 10.13040/IJPSR.0975-8232.12(1).330-35
CrossRef - Mahmud J., Khan R.A. Characterization of natural antimicrobials in food system. Adv Microb. 2018; 8: 894-916. DOI: 10.4236/aim.2018.811060
CrossRef - Rajkumar V., Berwal J.S. Inhibitory effect of clove (Eugenia caryophyllus) on toxigenic molds. J Food Sci Tech. 2003; 40: 416-418.
CrossRef - Kim H.O., Park S.W., Park H.D. Inactivation of Escherichia coli 0157:H7 by cinnamic aldehyde purified from Cinnamomum cassia shoot. J Food Microbiol 2004; 21: 105-110.
CrossRef - Hetta H.F., Meshaal A.K., Algammal A.M., Yahia R., Rabab R., Makharita R.R. In-vitro antimicrobial activity of essential oils and spices powder of some medicinal plants against Bacillus species isolated from raw and processed meat. Inf Drug Res. 2020;13: 4367–4378.DOI: 10.2147/IDR.S277295
CrossRef - Gutierrez-del-Rio I., Fernandez J., Lombo F. Plant nutraceuticals as antimicrobial agents in food preservation: terpenoids, polyphenols and thiols. Int J of Antimicrob Agents. 2018; 52:309-315. DOI: 10.1016/j.ijantimicag.2018.04.024
CrossRef - Papadochristopoulos A., Kerry J.P., Fegan N., Burgess C.M., Duffy G. Natural anti-microbials for enhanced microbial safety and shelf-life of processed packaged meat. Foods. 2021; 10: 01-42. DOI: 10.3390/foods10071598
CrossRef - Chouhan S., Sharma K., Guleria S. Antimicrobial activity of some essential oils-present status and future perspectives. Medicines. 2017; 4: 01-21. DOI:10.3390/medicines4030058
CrossRef - Sant D.G., Tupe S.G., Ramana C.V., Deshpande M.V. Fungal cell membrane-promising drug target for antifungal therapy. J Appl Microbiol. 2016; 121: 1498-1510.
CrossRef - Angane M., Swift S., Huang K., Butts C.A., Quek S.Y. Essential oils and their major components: an updated review on antimicrobial activities, mechanism of action and their potential application in the food industry. Foods. 2022; 11: 01-26. DOI: 10.3390/foods11030464
CrossRef - Haro-González J.N., Castillo-Herrera G.A., Martínez-Velázquez M., Espinosa-Andrews H. Clove essential oil (Syzygium aromaticum L. Myrtaceae) : Extraction, chemical composition, food applications, and essential bioactivity for human health. Molecules. 2021; 26: 01-25. DOI: 10.3390/molecules26216387
CrossRef - Chahbi A, Nassik S, Amri H, Douaik A, Maadoudi EHE, Boukharta M, Hadrami EME. Chemical composition and antimicrobial activity of the essential oils of two aromatic plants cultivated in Morocco (Cinnamomum cassia and Origanum compactum). J Chem. 2020; 2020: 01-10. DOI: 10.1155/2020/1628710
CrossRef - Dubey P.N., Saxena S.N., Lal G., Kant K., Mishra B.K., Agarwal D., John S. and Sharma L.K. Essential oil constituents and unsaturated fatty acids in Indian Cuminum cyminum L. seed oil under varying agro climatic environments. Int J seed spices. 2018; 8: 7-14.

This work is licensed under a Creative Commons Attribution 4.0 International License.






