
Introduction
Duck meat, which is commonly viewed as lower-value poultry meat, presents an exciting option as a potential source for surimi-like components. While most previous research on poultry-based surimi-like materials has focused on chicken, data are scarce on other fowl varieties such as duck, turkey, and quail. Non-chicken poultry meat has a high-fat level, heme pigment concentration, and collagen content, limiting its use as a surimi-like material source. Surimi technology, on the other hand, has gained popularity among non-fish animal species, particularly poultry.1
Duck flesh is high in polyunsaturated fatty acids, just like chicken meat.2 Duck flesh, on the other hand, is red, unlike chicken meat, due to the prevalence of red muscular fibers, particularly in the breast.3 Overfeeding considerably raises lipid levels in duck flesh, resulting in increased fat and moisture content in the breast muscle. In overfed ducks, this results in paler color, more yellowness, higher cooking loss values, less juiciness, and more pronounced flavor.4 Duck breast meat sensory quality is regulated by genotype and lipid levels, with increased lipid levels affecting tenderness, cooking loss, lightness, yellowness, and flavor.4
Duck flesh has the potential to be employed as a new form of surimi-like substance to boost its value. Underutilized duck meat can be rescued and turned into new products since it is richer in fat and darker in color.5 Surimi manufacturing is one method of reusing underutilized meat by washing the meat in various washing solutions.
Cryoprotectants are essential for maintaining the structural integrity of surimi proteins during freezing. Sucrose, sorbitol, and polyphosphates are popular cryoprotectants used in the manufacturing of fish surimi.6 However, the excessive sweetness of sucrose and sorbitol in the final product is a concern for consumers.7 Alternative cryoprotectants that have lower or absent sweetness for application in surimi manufacturing have been investigated by researchers. Palatinose, lactitol, and polydextrose tend to be less sweet than sucrose and sorbitol and have all been shown to stabilize fish surimi proteins during freezing.8
The objective of the current investigation was to assess the impact of low-sweetness cryoprotectants (PAL-Palatinose, TRE-Trehalose, PLD-Polydextrose, MAL-Maltodextrin, and LAC-Lactose) on the surimi-like material made from duck (DSLM) during frozen storage. Sucrose-sorbitol (SS) was also used for comparison with the low-sweetness cryoprotectants. The study aimed to identify the most effective cryoprotectants at preventing or slowing down freeze-induced alterations in duck muscle’s natural actomyosin (NAM) systems.
Materials and Methods
Raw Materials
Twenty-month-old native spent ducks (Khaki Campbell) with an average weighing 2 kg were purchased from a local farm. The slaughtering was conducted at the commercial slaughterhouse and the carcasses were deboned and frozen at -40°C overnight before being transported to the laboratory for further use.
Cryoprotectants
The cryoprotectants used in the manufacture of surimi-like material made from duck (DSLM) were obtained from both domestic and international suppliers. Crystalline LAC-lactitol monohydrate and powdered food-grade PLD-polydextrose (Danisco Sweeteners, Thomson, Illinois, USA), food-grade PAL-palatinose (Mannheim, Germany), MAL-maltodextrin (San Soon Seng Food Industries, Selangor, Malaysia), TRE-trehalose (treha, Hayashibara, Japan), crystalline food-grade sorbitol (Sorbogem™ 1162, crystalline, Euro Chemo-Pharma, Penang, Malaysia), and food-grade sodium tripolyphosphate (STPP, Euro Chemo-Pharma, Penang, Malaysia) were employed in this study.
Preparation of surimi-like material made from duck (DSLM)
The DSLM preparation was generated using the method outlined by Ismail et al.9. The frozen deboned meat was diced into small cubes using the chain saw (Hobart 6614, Troy, Ohio, USA) and ground into a meat mincer equipped with a standard hole diameter of 4.5 mm (Eve/All-12, Torino, Italy). Subsequently, the mince was weighed and subjected to two rounds of cold-water washing (4°C), at a ratio of 1:3 of meat to water (w/v). A universal mixer (B10-3, Selangor, Malaysia) was used to stir constantly for 4 min and was allowed to settle for 10 min. The settled residue was decanted after removing the topmost layer at 4000 rpm and 4°C for 15 min (Union 5 KR, Incheon, Korea). The liquid fraction was discarded, and the decanted meat was mixed with the following cryoprotectants:
The DSLM was divided into seven treatments by incorporating specific cryoprotectants:
LAC treatment: 6% lactitol and 0.3% sodium tripolyphosphate;
PLD treatment: 6% polydextrose and 0.3% sodium tripolyphosphate;
PAL treatment: 6% palatinose and 0.3% sodium tripolyphosphate;
MAL treatment: 6% maltodextrin and 0.3% sodium tripolyphosphate;
TRE treatment: 6% trehalose and 0.3% sodium tripolyphosphate;
SS treatment: 3% sorbitol, 3% sucrose, and 0.3% sodium tripolyphosphate;
Control (CON): DSLM with no cryoprotectant added.
The cryoprotectants were thoroughly mixed for 1 min using a high-speed mixer (Silent Cutter Hobart 84/45, Ohio, USA) at a chilled temperature. The DSLM was then frozen for 4 months at -18°C in sealed polyethylene packaging. Samples were collected for analysis at months 0, 1, 2, 3, and 4. Each experiment was conducted in triplicate.
Water Holding Capacity
The water retention ability was measured using a slightly modified method reported by Ramadhan et al.1 with a modification. To determine the water retention, a 20 g sample was blended using a waring blender with distilled water for 30 s to establish homogeneity before measuring the water retention. Next, 20 g of homogenate was carefully placed into a centrifuge tube and decanted at a low speed of 1000 g for 15 min while keeping the temperature at 5 °C. The water retention (WHC) was determined as liquid loss using the following formula:
WHC (%)= (Weight before centrifugation-Weight after ccentrifugation)/(Weight before centrifugation) ×100%
Extraction of Actomyosin
Actomyosin was extracted according to Nopianti et al.8. A 5 g sample was mixed for 4 min in 50 ml of cold 0.6 M potassium chloride at neutral pH with a homogenizer (Model IKA® T25, Germany). The sample was homogenized for 20 s and then given a 20 s resting period to prevent overheating during the extraction process. After that, the homogenate was subjected to centrifugation for 25 min at 4000 rpm at 0°C, followed by the addition of cold water (3 volumes) to precipitate the actomyosin. Centrifugation (4000 rpm) was performed under cold conditions (0°C) for 25 min. Next, the resulting pellet was dissolved by gentle stirring at a temperature <1°C for 25 min to the 1:1 ratio of pH 7.0 cold 1.2 M KCl. Any undissolved material was removed from the preparation by subjecting it to the same centrifugation conditions as mentioned above.
Myosin Ca2+-ATPase Activity
To analyze the oxidative deterioration of surimi protein after frozen storage, myosin Ca2+-ATPase activity was measured using the protocol outlined by Nurkhoeriyati et al.10. At pH 7.4, actomyosin (0.2 ml) was mixed with reaction solution of 2 ml containing 180 mM Tris-HCl, 15 mM CaCl2, 7.6 mM ATP, and 150 mM KCl. After letting the mixture react for 10 min at 25°C, the reaction was halted by adding 1 ml of trichloroacetic acid, 10%. Following the centrifugation of the combination for 5 min at 3500 rpm, the supernatant 1 ml was added with 3 ml of a solution containing 0.66% ammonium molybdate in 0.75 N sulfuric acid. To enhance color development, a freshly made 10% FeSO4 in 0.15 N sulfuric acid 0.5 ml aliquot was incorporated into the mixture, and the reaction was led to continue for 2 min. A sample blank comprising solely water was included in the analysis. The ATPase activity was determined by measuring the absorbance at 700 nm (Shimadzu UV-160A, Kyoto, Japan). A standard curve for phosphate calculation was prepared using NaH2PO4.
Total Sulfhydryl
The determination of total sulfhydryl (SH) was based on the method published by Nurkhoeriyati et al.10. 2 ml of urea-SDS solution (8 M urea, 3% SDS, 100 ml of phosphate buffer, pH 7.4) was added to 0.9 ml of diluted actomyosin. After that, the titration of SH was initiated by the addition of 1 ml of DTNB reagent, which was comprised of 10 mM DTNB in 0.1 M phosphate buffer with a pH of 7.4. The absorbance was measured at 412 nm using a spectrophotometer (UV-160A Shimadzu, Japan) after 15 minutes of incubation at room temperature. SH concentration was calculated using 11400 M-1cm-1 molar extinction.
Salt Extractable Protein (SEP)
SEP was determined using the Kjeldahl method.11 The proportion of soluble protein to total protein determined protein solubility.
(Total actomyosin protein)/(Total sample protein)×100%
Differential Scanning Calorimetry (DSC)
DSC was used to evaluate duck surimi actin and myosin temperature stability, as previously stated by Moosavi-Nasab et al.12. Thermal analysis of freeze-dried materials was performed using a Q100 differential scanning calorimeter equipped with a DSC refrigerated chilling system (Mettler DSC-30, Greinfensee, Switzerland). The analysis was tracked using the universal analysis 2000 software (ver.3.7A). 2 mg of the sample was put in aluminum pans and filled with phosphate buffer (pH 7.4) to 15 mg. The pans were then positioned on the Q100 calorimetric cell and completely sealed. The temperature of the cell was allowed to fall to 25°C before it was heated at a rate of 5°C per minute until it reached 100°C.
Scanning Electron Microscopy (SEM)
SEM analysis was performed on small samples with a diameter of 0.5 cm and a thickness of 0.3 cm.13 The samples were dehydrated overnight in a freeze drier. They were then carefully placed on aluminum stubs with adhesive and covered with 40-50 nm gold and measured by electron microscope (Philips 505, Eindhoven, The Netherlands).
Statistical Analysis
The trial was run in triplicate using SPSS version 23 (SPSS Inc., Chicago, Ill., USA). The obtained data were checked for normality using Levene’s test (P > 0.05), and the distribution of errors was determined using the Kolmogorov-Smirnov test (P > 0.05). The effect of cryoprotectant and storage was analyzed by the general linear model (P < 0.05). Multiple mean comparisons were performed using the Duncan test at a significance level of 5% if homogeneity of variance was not violated. In the event that this assumption was found to be violated, the Games-Howell test was conducted. Pearson correlation analysis was performed to establish a linear relationship between measurement.
Results and Discussion
Water Holding Capacity
Figure 1 presents the comparison of water holding capacity (WHC), measured as water loss, between DSLM samples with and without cryoprotectants. After 4 months in frozen storage, it was observed that the CON sample exhibited a significantly higher water loss percentage (52-56%) compared to the samples treated with cryoprotectants (37-47%). This discrepancy can be attributed to the degradation of myosin in the CON sample, which resulted in the formation of an inferior gel network. As a result, the gel matrix had decreased extensibility and a low WHC.14
The use of cryoprotectants has been proven beneficial in enhancing the gel-forming ability of DSLM. Walayat et al.6 observed comparable enhancements in prior investigations. The gel structure became more stable and resistant to water loss through the addition of cryoprotectants, keeping its ideal qualities. We also looked at the effects of cryoprotectants, storage, and the interaction between cryoprotectants and storage. The results demonstrate that there is a substantial effect on the WHC (P < 0.001). Several factors, including Ca2+-ATPase activity, SH concentration, and protein solubility, were examined to highlight the CON samples’ insufficient water binding capacity. These data provided additional evidence that myosin content decreased during storage, showing protein breakdown over time, which directly weakened the gel matrix’s efficient water retention ability.
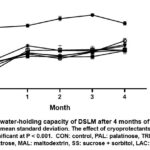 |
Figure 1: Mean water-holding capacity of DSLM after 4 months of frozen storage. |
Myosin Ca2+-ATPase Activity
Ca2+-ATPase activity is employed to determine the structural integrity of myofibrillar proteins as well as the degree to which they have been denatured when surimi is being frozen for storage. This activity represents the effects of denatured proteins and protein aggregation, leading to structural changes in myosin and decreased ATPase activity.15 Ca2+-ATPase activity in DSLM samples containing various cryoprotectants varied throughout frozen storage (Figure 2). Ca2+-ATPase activity decreased significantly in the SS (70.0%), LAC (65.1%), PLD (64.8%), MAL (58.5%), and TRE surimi (50.6%) after the initial month of frozen storage. This decrease showed that the freezing and thawing processes were causing myosin to be disrupted. When the activities of the CON samples and those with cryoprotectants were compared, it was clear that all cryoprotectants had a strong cryoprotective impact during the freezing process (Figure 2). The Ca2+-ATPase activity of the CON samples dropped considerably over time, eventually decreasing to undetectable levels after 3 months of frozen storage.
Cryoprotectants play a crucial role in preserving actomyosin by stabilizing its structure.6 They keep the protein from losing water, increase the surface tension of water7, enhance the amount of bound water, and prevent the loss of protein solubility during frozen storage.6 Freezing commonly damages muscle protein, leading to protein denaturation, loss of functionality16, reduced solubility of myofibrillar protein, disappearance of ATP-induced muscle fiber contraction, and decreased myosin ATPase activity.17 The degradative effect is evident both with and without cryoprotectant, as depicted in Figure 2. Nonetheless, the treated samples with cryoprotectant demonstrate a milder impact compared to the CON sample. This is because protein denaturation takes place in the absence of cryoprotectant.
According to the findings of this investigation, PAL effectively avoided a substantial decrease in Ca2+-ATPase activity throughout the first month of storage. Although PAL initially showed lower activity compared to other cryoprotectants at month 0 (Figure 2), it consistently decreased to the lowest at 3 months of storage among the cryoprotectant-added samples. We empirically observed that PLD had greater Ca2+-ATPase activity stability over the whole frozen storage period. The presence of PLD appeared to have a protective impact on myosin, protecting it from the negative effects of freezing-induced microenvironmental alterations. Throughout the 4-month frozen storage period, these alterations damaged myosin progressively, albeit to a significantly smaller amount in the PLD samples compared to the other cryoprotectant-added samples. Polydextrose functions as an effective anti-freezing agent and stabilizer. When 8% polydextrose was incorporated into surimi, it exhibited cryoprotective properties.6 Furthermore, polydextrose is an effective cryoprotectant due to its great water solubility, high viscosity, and hygroscopic nature, making it appropriate for long-term preservation.18
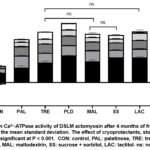 |
Figure 2: Mean Ca2+-ATPase activity of DSLM actomyosin after 4 months of frozen storage. |
Changes in Total Sulfhydryl Content
Sulfhydryl (SH) values were significantly affected by cryoprotectant (Pc < 0.01), storage (Ps < 0.001) and their interaction (Pc × s < 0.001). These observations show that the samples treated with cryoprotectant are more effective in slowing the sulfhydryl loss during frozen storage. SH groups in DSLM must exist for the development of gel networks and the binding of meat particles. The SH group at the head position plays a vital role in modulating Ca2+-ATPase activity. Notably, this SH content demonstrates a significant positive correlation (r2 = 0.591) with Ca2+-ATPase activity, exhibiting a concurrent decrease as Ca2+-ATPase activity decreases.15 Throughout the frozen storage period (Table 1), the SH content of DSLM exhibited a consistent reduction. The decline in SH content from months 0 to 4 was observed in various samples, including CON, PAL, TRE, PLD, MAL, SS, and LAC, with values ranging from 26.27 to 8.58, 24.10 to 15.89, 28.09 to 11.65, 27.51 to 13.47, 26.24 to 10.09, 35.94 to 8.05, and 23.66 to 10.00 nanomol SH/mg protein, respectively. During the frozen storage of surimi, the SH groups undergo oxidation, forming disulfide bonds that facilitate cross-linking among protein molecules. Consequently, the SH content diminishes.19 The CON sample exhibited the most substantial loss, which can be attributed to the absence of a cryoprotectant that would otherwise slow down myofibrillar protein conformational changes during frozen storage.
The SH content of natural actomyosin from surimi is particularly susceptible to oxidation. Approximately 65% of the SH from myofibril preparations is located in the myosin molecule, implying that myosin oxidation may be the cause of an important portion of the SH loss resulting from freeze-thaw stress.8 Table 1 illustrates that none of the cryoprotectants could prevent SH oxidation in DSLM during frozen storage. Therefore, freezing, thawing, and long-term storage can all cause SH depletion. In this study, the oxidation of SH was slowed down in DSLM with PAL and PLD added, indicating that these cryoprotectants had a protective effect by delaying the rate of SH oxidation.
Table 1: Variation in the total number of sulfhydryl in actomyosin with different cryoprotectants during 4-month storage
|
Storage (month) |
CON (nanomol SH/mg protein) |
PAL (nanomol SH/mg protein) |
TRE (nanomol SH/mg protein) |
PLD (nanomol SH/mg protein) |
MAL (nanomol SH/mg protein) |
SS (nanomol SH/mg protein) |
LAC (nanomol SH/mg protein) |
Ps |
|
0 |
26.27 |
24.10 |
28.09 |
27.51 |
26.24 |
35.94 |
23.66 |
P < 0.001 |
|
1 |
19.38 |
20.21 |
17.20 |
18.99 |
24.21 |
20.51 |
21.16 |
P < 0.001 |
|
2 |
14.97 |
20.19 |
16.48 |
18.11 |
17.75 |
19.79 |
19.97 |
P < 0.001 |
|
3 |
13.78 |
20.13 |
14.36 |
16.50 |
14.82 |
10.41 |
11.91 |
P < 0.001 |
|
4 |
8.58 |
15.89 |
11.65 |
13.47 |
10.09 |
8.05 |
10.00 |
P < 0.001 |
|
Pc |
P |
P |
P |
P |
P |
P < 0.001 |
P |
|
|
Pc × Ps |
P |
P |
P |
P |
P |
P |
P |
|
Each value is expressed as mean ± standard deviation
Different lowercase and uppercase letters within the same column and row represent statistically significant differences (P < 0.05).
Pc: significant effect of cryoprotectants; Ps: significant effect of storage; Pc * Ps: significant interaction between cryoprotectant and storage
CON: control, PAL: palatinose, TRE: trehalose, PLD: polydextrose, MAL: maltodextrin, SS: sucrose + sorbitol, LAC: lactitol.
Changes in Protein Solubility
As shown in Table 2, the protein solubility data collected from the various samples during the various storage periods provide useful insights into the effects of frozen storage and the possible impact of cryoprotectants on protein stability. Protein solubility decreases are a prominent indicator of freeze-induced protein denaturation. Denaturation occurs during the frozen storage process due to the production of hydrogen bonding, as well as the formation of sulfide bonds and ionic interactions.15 The findings reveal a general trend of reduced protein solubility as storage time increases. This decline may be observed in all samples, suggesting that proteins denature over time under frozen storage conditions.
Interestingly, some cryoprotectants appear to affect protein solubility. At each storage time, PAL had relatively higher protein solubility than the other samples among the cryoprotectants tested. This suggests that PAL, as a cryoprotectant, may have a protective impact on protein structure, assisting in the preservation of solubility during frozen storage. It is important to note that protein solubility declines over time, albeit at a somewhat slower rate than in the absence of cryoprotectants. When we compare the changes in cryoprotectants after 4 months, we notice varied amounts of protein solubility. PLD, TRE, and SS, for example, show a lesser drop in solubility when compared to PAL and other cryoprotectants. The solubility loss values for these cryoprotectants are less than 40% compared to the initial level at 0 months. In contrast, MAL and LAC show substantial reductions in protein solubility, reaching 18.60% and 16.32%, respectively, during the 4-month storage period, which is lower than the CON sample. These results imply that maltodextrin and lactitol, as cryoprotectants, may not be as effective in preserving protein solubility during frozen storage compared to other cryoprotectants evaluated in this study. The statistical study also revealed a substantial interaction between storage duration and cryoprotectants, demonstrating that both factors influenced protein solubility (Pstorage * cryoprotectants: P < 0.001).
During the freezing storage period, protein solubility undergoes a decline as a result of multiple factors, including ice crystal formation, protein aggregation, and structural changes.20 These variables disturb the native structure of proteins, exposing hydrophobic areas and forming interactions between molecules, which ultimately results in reduced solubility.21 Previous research has shown that integrating cryoprotectants can effectively sustain increased protein solubility over 4 months when compared to samples without cryoprotectant additions.18 Additionally, Zhou et al.22 observed that 8% (w/w) trehalose and sodium lactate efficiently reduced the denaturation of proteins in tilapia surimi stored in the freezer for 24 weeks at -18°C. Moreover, Nopianti et al.8 found that polydextrose exhibited the highest protein solubility values among the tested threadfin bream samples throughout the six-month frozen storage period.
Table 2: Variation in protein solubility in actomyosin with different cryoprotectants during 4-month storage
|
Storage (month) |
CON (%) |
PAL (%) |
TRE (%) |
PLD (%) |
MAL (%) |
SS (%) |
LAC (%) |
Ps |
|
0 |
56.82 |
46.25 |
37.14 |
45.10 |
40.01 |
39.18 |
39.68 |
P = 0.005 |
|
1 |
45.25 |
50.40 |
45.00 |
46.45 |
43.28 |
49.54 |
46.08 |
P = 0.377 |
|
2 |
47.51 |
52.98 |
47.70 |
53.64 |
46.28 |
36.22 |
51.92 |
P = 0.012 |
|
3 |
33.62 |
37.15 |
33.31 |
43.27 |
30.71 |
34.74 |
32.70 |
P = 0.020 |
|
4 |
29.40 |
29.98 |
30.34 |
35.43 |
18.60 |
31.51 |
16.32 |
P < 0.001 |
|
Pc |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P |
P < 0.001 |
P = 0.003 |
P = 0.001 |
|
|
Pc × Ps |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P |
P < 0.001 |
P < 0.001 |
P < 0.001 |
Each value is expressed as mean ± standard deviation
Different lowercase and uppercase letters within the same column and row represent statistically significant differences (P < 0.05).
Pc: significant effect of cryoprotectants; Ps: significant effect of storage; Pc * Ps: significant interaction between cryoprotectant and storage
CON: control, PAL: palatinose, TRE: trehalose, PLD: polydextrose, MAL: maltodextrin, SS: sucrose + sorbitol, LAC: lactitol.
Differential Scanning Calorimetry
DSC is an effective method for determining how temperature affects the specific heat of food samples. It offers a rapid and simple method, providing accurate results even with small sample sizes.23 In the present study, DSC analysis was conducted on myofibrillar proteins derived from DSLM with various cryoprotectants during 4-month storage. The thermograms obtained from the DSC analysis exhibited two distinct endothermic peaks corresponding to myosin and actin (Figure 3a). However, over time (by 4-month storage), these endothermic peaks, were gradually decreased in intensity.
Differences in the transitions of myosin and actin at different storage periods are illustrated using box plots in Figure 4. Each box on the graphic depicts the interquartile temperature range (°C), which encompasses the center 50% of data points. The median is represented by the horizontal line inside the box, while the mean is represented by the cross. The whiskers (two lines extending from the box) show the extent of data distribution. Outliers are indicated by dot points outside the whiskers. For myosin endothermic value, the box plots demonstrate that there was significant overlap at 0-month storage between the samples with and without cryoprotectant, except for the TRE sample (P > 0.05). Interestingly, PLD showed a more stable temperature transition compared to the other samples after 4 months of frozen storage. The largest decrease in effectiveness was recorded for LAC (10.8%), TRE (9.7%), and CON (7.0%) compared to the initial values at 0 months. On the other hand, the smallest decrease was observed in PLD (2.8%), followed by SS (4.7%), PAL (5.1%), and MAL (5.8%). These findings suggest that the absence of cryoprotectant (CON) and the addition of LAC and TRE were not suitable for producing desirable surimi due to a higher percentage of myosin loss during prolonged frozen storage. This indicates that these samples caused protein structure destabilization during storage. As the storage time increased, the thermal curves began to change, and by month 4, the transition of myosin became less distinct (Figure 3b). Indeed, the degree of protein denaturation increases gradually over time (P < 0.001), and the influence of the cryoprotectant is also evident (P < 0.01) through the observation of denaturation enthalpies (∆H) as shown in Table 3. Notably, myosin exhibits a significant reduction in ∆H when treated with MAL, LAC, TRE, and CON after 4 months of storage. The most substantial ∆H value for myosin is achieved in the samples blended with SS and PAL at the initial month of storage. Empirically, the stability across a 4-month frozen storage period with minimal denaturation of myosin can be ranked as follows: PAL > PLD > SS > MAL > TRE > LAC > CON, with denaturation percentages of 13%, 39%, 55%, 56%, 71%, 81%, and 90%, respectively. The magnitude of the ∆H value relies on the concentration of folded protein fractions. If the initial folded protein fraction is less than 100% of the total protein concentration, the calculated ∆H value will proportionally decrease. As noted by Kovacevic et al.,24 higher ∆H values correlate with a greater cryoprotective effect of the cryoprotectant.
For actin, the enthalpy of denaturation also decreased but remained relatively stable during the 0 to 4-month storage period, except for CON, where a higher denaturation rate of 69% was observed. In contrast, samples treated with the cryoprotectant exhibited improved ∆H values for actin after 4 months of storage. It is worth noting that myosin undergoes a higher level of degradation compared to actin, as myosin denatures at temperatures lower than those required for actin denaturation. Specifically, myosin denatures at 60°C, while actin denatures within the temperature range of 66-73°C.25 This observation emphasizes the significance of myosin as a critical functional protein in the formation of surimi gel.26
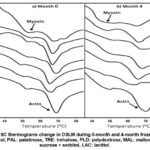 |
Figure 3: DSC thermograms change in DSLM during 0-month and 4-month frozen storage. |
 |
Figure 4: DSC thermograms change (Tp) in DSLM during 0 and 4-month frozen storage. |
Table 3: Denaturation enthalpies (∆H) of myosin and actin with different cryoprotectants during 0 and 4-month storage
|
Storage (month) |
CON ∆H (J/g) |
PAL ∆H (J/g) |
TRE ∆H (J/g) |
PLD ∆H (J/g) |
MAL ∆H (J/g) |
SS ∆H (J/g) |
LAC ∆H (J/g) |
Ps |
|
Myosin |
|
|
|
|
|
|
||
|
0 |
0.30 |
0.84 |
0.69 |
0.51 |
0.11 |
0.89 |
0.43 |
P < 0.001 |
|
4 |
0.03 |
0.73 |
0.20 |
0.31 |
0.05 |
0.40 |
0.08 |
P < 0.001 |
|
Pc |
P |
P |
P |
P |
P |
P |
P |
|
|
Pc × Ps |
P |
P |
P |
P |
P |
P |
P |
|
|
Actin |
||||||||
|
0 |
0.39 |
1.08 |
0.72 |
1.04 |
0.79 |
0.79 |
0.82 |
P < 0.001 |
|
4 |
0.12 |
0.76 |
0.39 |
0.74 |
0.49 |
0.51 |
0.48 |
P < 0.001 |
|
Pc |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
|
|
Pc × Ps |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
P < 0.001 |
Each value is expressed as mean ± standard deviation
Different uppercase letters within the same row represent statistically significant differences (P < 0.05).
Pc: significant effect of cryoprotectants; Ps: significant effect of storage; Pc * Ps: significant interaction between cryoprotectant and storage
CON: control, PAL: palatinose, TRE: trehalose, PLD: polydextrose, MAL: maltodextrin, SS: sucrose + sorbitol, LAC: lactitol.
Scanning Electron Microscopy (SEM)
SEM is a valuable technique utilized for evaluating the morphology of surimi gels, notably the size and distribution of micro-pores on the surface. Surimi gel’s physicochemical qualities are heavily influenced by its microstructure. In the absence of cryoprotectants (Figure 5A), surimi gels tend to undergo an enlargement of pore sizes during frozen storage, resulting in a loss in WHC and textural features. However, when treated with cryoprotectants, the surimi gel samples exhibited a compact structure and demonstrated minimal changes in pore size, except for the LAC-treated samples (Figure 5B). After 4 months of storage, the LAC-treated samples had a higher myosin transition temperature, a more significant drop in protein solubility (69%), and poorer Ca2+-ATPase activity. Although there were no significant alterations in WHC, these findings were observed directly through the microstructure, showing that LAC was destabilized.
In comparison to other samples, DSLM prepared with PLD and PAL (Figure 5D and F, respectively) exhibited a compact and dense microstructure, whereas the remaining samples displayed a coarser and disordered gel structure with clusters of agglomerates. This shows that the cryoprotectants used aided in the creation of a well-structured gel matrix. The cryoprotectants promoted the homogeneous cross-linking of myofibrillar proteins and water molecules, as a result, the gel structure exhibits a higher specific surface area and an increased interaction between them.27 Consequently, DSLM with cryoprotectants exhibited a higher WHC, as the water molecules did not easily migrate, underscoring the close relationship between the microstructure and WHC of the gels.
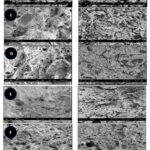 |
Figure 5: Scanning electron micrographs of DSLM with and without cryoprotectants at month 0 and month 4. |
Conclusion
In conclusion, the addition of cryoprotectants to surimi-like material made from duck (DSLM) showed positive effects on its freeze-thaw stability. The interaction between cryoprotectant and storage had a significant effect on water holding capacity (WHC), sulfhydryl (SH) content, Ca2+-ATPase activity, and protein solubility. Among the cryoprotectants studied, polydextrose (PLD) emerged as the most effective in preserving Ca2+– ATPase activity, SH content, protein solubility, myosin transition, and microstructure of DSLM during 4 months of frozen storage, followed by PAL. Both PLD and PAL can serve as viable alternatives to sucrose sorbitol (SS). However, cryoprotectants such as MAL, LAC, and TRE showed some level of effectiveness but were not as reliable in improving the freeze-thaw stability of DSLM over prolonged storage.
Acknowledgement
The author would like to thank, Universiti Sultan Zainal Abidin, Besut Campus, Terengganu, Malaysia for their guidance and support to complete this article.
Conflict of Interest
The authors do not have any conflict of interest.
Funding Sources
The author(s) received no financial support for the research, authorship, and/or publication of this article.
References
- Ramadhan K., Huda N., Ahmad R. Effect of Number and Washing Solutions on Functional Properties of Surimi-like Material from Duck Meat. J Food Sci Technol. 2014;51(2):256-266. https://doi.org/10.1007/s13197-011-0510-1.
CrossRef - Chang Y.-S., Chen J.-W., Wu Y.-H. S., Wang S.-Y., Chen Y.-C. A Possible Systematic Culinary Approach for Spent Duck Meat: Sous-Vide Cuisine and its Optimal Cooking Condition. Poult Sci. 2023;102(6):102636. https://doi.org/10.1016/j.psj.2023.102636.
CrossRef - Ismail I., Joo S. T. Poultry Meat Quality in Relation to Muscle Growth and Muscle Fiber Characteristics. Korean J Food Sci Anim Resour. 2017;37(6):873-883. https://doi.org/10.5851/kosfa.2017.37.6.87.
- Chartrin P., Méteau K., Juin H., Bernadet M. D., Guy G., Larzul C., Rémignon H., Mourot J., Duclos M. J., Baéza E. Effects of Intramuscular Fat Levels on Sensory Characteristics of Duck Breast Meat. Poult Sci. 2006;85(5):914-922. https://doi.org/10.1093/ps/85.5.914.
CrossRef - Biswas S., Banerjee R., Bhattacharyya D., Patra G., Das A. K., Das S. K. Technological Investigation into Duck Meat and its Products – A Potential Alternative to Chicken. Worlds Poult Sci J. 2019;75(4):609-620. https://doi.org/10.1017/S004393391900062X.
CrossRef - Walayat N., Xiong H., Xiong Z., Moreno H. M., Nawaz A., Niaz N., Randhawa M. A. Role of Cryoprotectants in Surimi and Factors Affecting Surimi Gel Properties: A Review. Food Rev Int. 2022;38(6):1103-1122. https://doi.org/10.1080/87559129.2020.1768403.
CrossRef - Nikoo M., Benjakul S. Potential Application of Seafood-derived Peptides as Bifunctional Ingredients, Antioxidant–Cryoprotectant: A review. J Func Foods. 2015;19:753-764. https://doi.org/10.1016/ j.jff.2015.10.014.
CrossRef - Nopianti R., Huda N., Noryati I., Fazilah A., Easa A. M. Cryoprotective Effect of Low-sweetness Additives on Protein Denaturation of Threadfin Bream Surimi (Nemipterus Spp.) During Frozen Storage. CYTA J Food. 2012;10(3):243-250. https://doi.org/10.1080/ 19476337.2011.639033.
CrossRef - Ismail I., Huda N., Ariffin F., Ismail N. Effects of Washing on the Functional Properties of Duck Meat. Int J Poult Sci. 2010;9(6):556-561. https://doi.org/10.3923/ijps.2010.556.561.
CrossRef - Nurkhoeriyati T., Huda N., Ahmad R. Gelation Properties of Spent Duck Meat Surimi-like Material Produced using Acid–Alkaline Solubilization Methods. J Food Sci. 2011;76(1):S48-S55. https://doi.org/10.1111/j.1750-3841.2010.01963.x.
CrossRef - AOAC. Official methods of analysis. 16th ed., Arlington: Association of Official Analytical Chemists; 2002.
- Moosavi-Nasab M., Alli I., Ismail A., Ngadi M. Protein Structural Changes During Preparation and Storage of Surimi. J Food Sci. 2005;70(7):c448-c453. https://doi.org/10.1111/j.1365-2621.2005.tb11467.x.
CrossRef - Andrés S. C., Garcı́a M. E., Zaritzky N. E., Califano A. N. Storage Stability of Low-fat Chicken Sausages. J Food Eng. 2006;72(4):311-319. https://doi.org/10.1016/j.jfoodeng.2004.08.043.
CrossRef - Tang S., Feng G., Cui W., Gao R., Bai F., Wang J., Zhao Y., Zeng M. Effect of α-Tocopherol on the Physicochemical Properties of Sturgeon Surimi during Frozen Storage. Molecules. 2019;24(4):710. https://doi.org/10.3390/molecules24040710.
CrossRef - Zhang X., Zhang Y., Dong Y., Ding H., Chen K., Lu T., Dai Z. Study on the Mechanism of Protein Hydrolysate Delaying Quality Deterioration of Frozen Surimi. LWT. 2022;167:113767. https://doi.org/ 10.1016/j.lwt.2022.113767.
CrossRef - Tan M., Ding Z., Xie J. Freezing-induced Myofibrillar Protein Denaturation: Contributions of Freeze-concentration and Role of Cellobiose. J Food Eng. 2022;329:111076. https://doi.org/10.1016/ j.jfoodeng.2022.111076.
CrossRef - Zhang B., Hao G.-J., Cao H.-J., Tang H., Zhang Y.-Y., Deng S.-G. The Cryoprotectant Effect of Xylooligosaccharides on Denaturation of Peeled Shrimp (Litopenaeus Vannamei) Protein during Frozen Storage. Food Hydrocoll. 2018;77:228-237. https://doi.org/10.1016/ j.foodhyd.2017.09.038.
CrossRef - Ramadhan K., Huda N., Ahmad R. Freeze-thaw Stability of Duck Surimi-like Materials with Different Cryoprotectants Added. Poult Sci. 2012;91(7):1703-1708. https://doi.org/10.3382/ps.2011-01926.
CrossRef - Wang Z., He Z., Gan X., Li H. Effect of Peroxyl Radicals on the Structure and Gel Properties of Isolated Rabbit Meat Myofibrillar Proteins. IJFST. 2018;53(12):2687-2696. https://doi.org/10.1111/ijfs.13878.
CrossRef - Walayat N., Wang X., Nawaz A., Zhang Z., Abdullah, Khalifa I., Saleem M. H., Mushtaq B. S., Pateiro M., Lorenzo J. M., Fiaz S., Ali S. Ovalbumin and Kappa-Carrageenan Mixture Suppresses the Oxidative and Structural Changes in the Myofibrillar Proteins of Grass Carp (Ctenopharyngodon idella) during Frozen Storage. Antioxidants. 2021;10(8):1186. https://doi.org/10.3390/antiox10081186.
CrossRef - Han J., Sun Y., Zhang T., Wang C., Xiong L., Ma Y., Zhu Y., Gao R., Wang L., Jiang N. The Preservable Effects of Ultrasound-Assisted Alginate Oligosaccharide Soaking on Cooked Crayfish Subjected to Freeze-thaw Cycles. Ultrasonics Sonochemistry. 2023;92:106259. https://doi.org/ 10.1016/j.ultsonch.2022.106259.
CrossRef - Zhou A., Benjakul S., Pan K., Gong J., Liu X. Cryoprotective Effects of Trehalose and Sodium Lactate on Tilapia (Sarotherodon Nilotica) Surimi during Frozen Storage. Food Chem. 2006;96(1):96-103. https://doi.org/10.1016/j.foodchem.2005.02.013.
CrossRef - Zhang J., Farkas B. E., Hale S. A. Thermal Properties of Skipjack Tuna (Katsuwonus Pelamis). Int J Food Prop. 2001;4(1):81-90. https://doi.org/10.1081/JFP-100000345.
CrossRef - Kovacevic D., Mastanjevic K., Kordic J. Cryoprotective Effect of Polydextrose on Chicken Surimi. Czech J. Food Sci. 2011;29(3):226-231. https://doi.org/10.17221/201/2008-CJFS.
CrossRef - Ismail I., Hwang Y.-H., Joo S.-T. Interventions of Two-stage Thermal Sous-vide Cooking on the Toughness of Beef Semitendinosus. Meat Sci. 2019;157:107882. https://doi.org/10.1016/j.meatsci.2019.107882.
CrossRef - Gao X., Yang S., You J., Yin T., Xiong S., Liu R. Changes in Gelation Properties of Silver Carp Myosin Treated by Combination of High-Intensity Ultrasound and NaCl. Foods. 2022;11(23):3830. https://doi.org/10.3390/foods11233830.
CrossRef - Le H., Ting L., Jun C., Weng W. Gelling Properties of Myofibrillar Protein from Abalone (Haliotis Discus Hannai Ino) Muscle. Int J Food Prop. 2018;21(1):277-288. https://doi.org/10.1080/ 10942912.2018.1454463.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








