
Introduction
Andaliman (Zanthoxylum acanthopodium DC.) has been used as a food condiment in traditional cuisine for centuries in Batak ethnic in North Sumatra, not only because of its unique flavor, but it is also known to have folkloric usage against diarrhea, stomach ache, toothache, and used as the food preservative. The plant is a low branching shrub grown at an elevation of 900 m above sea level endemically around Lake Toba, North Sumatra1. Andaliman belongs to the citrus family (Rutaceae), which fruits have a diameter of 2-3 mm. In the native regions, it is also known as Sinyar-Sinyar, Syarnyar, Toba, or Intir-Intir. Previously, three varieties of this spice existed, namely Simanuk, Sihorbo, and Sitanga2; however, Simanuk was more commonly used due to its more desirable flavor and higher production yield. Sitanga is no longer found naturally. The fruits and leaves from similar plants, i.e., Z. piperitum and Z. simulans grown in China, Japan, and Korea, have also been used as food condiments and medicinal plants. Zanthoxylum species had been reported to have bioactivity toward the gastrointestinal tract3–6. Sanshools were known as one of main components that contributed to this bioactivity3,4.
Andaliman possessed several bioactivities such as antioxidant and antidiabetic7,8, anti-inflammatory9, the cytotoxic effect on breast cancer cells10,11, antihyperglycemic12, and anti-aging as well as antiacne effects13. However, no scientific study has reported its health benefit on the gastrointestinal tract nor bioactive compound. Considering its traditional value and its reported health benefit, Andaliman has a potency to be developed as a functional food ingredient to prevent degenerative diseases, especially in the gastrointestinal tract.
Colorectal cancer (CRC) has become a global health burden, with constantly increasing occurrence. The latest database in 2018 from the International Agency for Research in Cancer14 indicated that CRC was the third-highest cancer incidence (1,849,518 cases) and the second-highest cancer mortality (880,792 deaths) in the world. Meanwhile in Indonesia, CRC was the sixth-highest cancer incidence (17,368 cases) and the eighth-highest cancer mortality (9,444 deaths)15.
The development of CRC can be caused by three main pathways, i.e. chromosomal instability (CIN), microsatellite instability (MSI), and CpG island methylator phenotype (CIMP). For instance, DNA hypomethylation and hypermethylation in the specific region result in the inactivation of tumor suppressor16. Our study address the role of apoptotic event to regulate cells proliferation. Fas protein (an apoptotic factor) is known to affect the chromosomal stability via tumor suppressor transcription factor 53 (TP53) pathway16. Controlling FasR/FasL signaling pathway17 has been reported as one of the methods studied in the preclinical models for colon cancer cells. The other studies also have shown the increase tumor medicinal agent efficacy with the addition of methyl-β-cyclodextrin, as a cholesterol suppressor. This heptasaccharide was able to promote apoptosis and PARP-1 cleavage through p53 activation to inhibit MCF-7 cell, Hepa 1-6 cell, and melanoma cancer cells development18,19.
Besides targeted clinical treatments, cancer chemoprevention has recently become a new approach to reduce the occurrence and burden of CRC. The chemopreventive intervention, such as dietary agents and pharmaceuticals, could help to inhibit, reverse, or delay the growth of abnormal cells into neoplastic diseases20, in addition to the induction of apoptosis21. In supporting this approach, many scientists have done research related to natural chemopreventive agents from herbs and spices22,23. Therefore, this research aimed to evaluate the cytotoxicity induced by Andaliman extract toward human colon cancer cells and the underlying mechanism of action in the colon cancer cells inhibition.
Materials and methods
Plant material and sample preparation
Andaliman (Z. acanthopodium DC. var. Simanuk) fruits were collected from a farmer in Dairi Regency, North Sumatera, Indonesia. The fruits used in this experiment were green and fresh (2-3 days old from the harvest day). To preserve the sample, it was oven-dried at 54 °C for eight hours before the extraction24. The plants were identified by “Herbarium Bogorience” Research Center for Biology in the Indonesian Institute of Sciences (LIPI) with specimen No. 382/IPH.1.01/If.07/11/2017.
Extraction
One hundred grams of Andaliman fruits were crushed for 10-15 s and were macerated for 72 hours twice at room temperature using 220 mL analytical-grade ethanol (Merck, Germany). It was then filtered using filter paper (Whatman no.1). The filtrate was evaporated under low pressure at a temperature of 40 °C using a rotary vacuum evaporator (Büchi Rotavapor® R-11) for ± 90 mins (pressure: 1000 mbar; rotary scale: 3). Finally, the crude extract was freeze-dried using a laboratory-scale benchtop freeze drier (Martin Christ Alpha 1-2 LDplus).
Cell culture maintenance
WiDr (ATCC® CCL-218™) and HCT-116 (ATCC® CCL-247™) are human colon cancer cells obtained from Primate Center Research, IPB University. WiDr and HCT-116 cells were between passages 17-21 and 32-36, respectively, for all the experiments. WiDr cells were cultured in RPMI (Roswell Park Memorial Institute) 1640 Medium; meanwhile, HCT-116 were cultured in DMEM (Dulbecco’s Modified Eagle Medium). Both of the media were supplemented with 10% (v/v) fetal bovine serum (FBS) (Gibco, Life Technologies Corporation, USA) and 100 U.ml-1 penicillin/streptomycin (Gibco, Life Technologies Corporation, USA). Cells were incubated at 37 °C and 5% CO2. The growth media was changed every two days. Subculture was done by washing the cells using PBS (phosphate-buffered saline) and adding 0.5 mL of 0.25% trypsin. After incubation for 5 mins, 1 mL of media was added to inactivate the trypsin. The suspension was centrifuged at 250xg for five mins. The cell pellet was resuspended with 3 mL media and counted. An estimated 5×104 cells.mL-1 was removed to a T25 flask and 10 mL of media was added.
Cytotoxicity analysis
Cell culture was transferred to a 96-wells microplate, 100 µl for each well with a density of 5 x 103 cells/well and three wells were left as the media control. The condition and distribution of the cells were observed using an inverted microscope. After an overnight incubation, the media was discarded, and 100 µl of PBS was added and then thrown away. The extracts of Andaliman fruit with different concentrations were pipetted into the wells and incubated in a CO2 incubator for 48 hours. Andaliman extract was applied to WiDr and HCT-116 cells at concentrations of 213.33, 106.67, 53.33, 26.67, 13.33, and 6.67 μg.mL-1. Control cells were incubated without extract. Doxorubicin (Dox) 1 and 3 μg.mL-1 were also applied to both cells as the positive controls. Cytotoxicity analysis was done with MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide tetrazole) analysis to measure the inhibition of cells proliferation by the extracts. Tetrazolium salt reagent (10 μL; 5 mg.mL-1) was added to each well and was incubated for four hours; then, 96% ethanol was added to dissolve the reagent. The viability of colon cancer cells was evaluated using the iMark™ microplate absorbance reader (Bio-Rad Laboratories, Inc., UK) at 595 nm. All the treatments were performed in triplicates. The IC50 was calculated using the regression equation of the percentage of the colon cancer cells inhibition.
Bcl-2 and Bax gene expression analysis
Based on the results of cytotoxicity analysis, three concentrations of Andaliman extracts were applied to WiDr and HCT-116 cells, namely, 2, 1, and ½ times of the IC50. Both cells were transferred to a 6-wells microplate, 1 mL for each well with a density of 1.0 x 104 cells/mL. After 48 hours of treatment with the extract, the cells were harvested for RNA isolation. Untreated cells were used as a negative control.
Total RNA was isolated using RNA Mini Kit (50) Qiagen®. Every sample was diluted in 100 μL of RNAse-free water to detach the RNA. The quantity of RNA was measured by Nanodrop 2000C UV Spectrophotometer. Complementary DNA (cDNA) was synthesized using Primer Invitrogen SuperScript III First-Strand Synthesis System for RT-PCR (SS III RT) Kit following the instructions for use. The reactants were incubated at 25 °C for 10 mins, 50 °C for 59 mins, 85 °C for 5 mins, and 4 °C for 2 mins. RNAse H enzyme was added to harvest the cDNA. The reaction was done at 37 °C for 20 mins and 4 °C until done. The RT-PCR analysis to measure the expression of Bcl-2 and Bax was carried out using SSo fast evergreen Supermix and performed using iQ5 Bio-Rad Real-Time PCR. Primer accession numbers, sequences (confirmed with BLAST analysis: www.ncbi.nlm.nih.gov/blast), and the annealing temperature (Ta) are shown in Table 1. The relative expression of both genes was analyzed using the 2-ΔΔCt method25. Each experiment was performed in duplicate. β-actin (ACTB) was used to normalize the gene expression levels.
Table 1: Primer sequences of target and normalization genes in WiDr and HCT-116 cells.
| Gene | Accession numbers | Primer sequences | Ta (°C) |
| Bcl-226 | NG – 009361.1 | F: 5′- GCTCTAAAATCCATCCAG -3′R: 5′- CCTCTCCATCATCAACTT -3′ | 48 |
| Bax27 | NG – 012191.1 | F: 5′- CCCGAGAGGTCTTTTTCCGAG -3′R: 5′- CCAGCCCATGATGGTTCTGAT -3′ | 67 |
| β-actin27 | NG – 007992.1 | F: 5′- AGAGCTACGAGCTGCCTGAC -3′R: 5′- AGCACTGTGTTGGCGTACAG -3′ | 67 |
*Ta: annealing temperature
Hoechst staining
To further confirm the apoptosis mechanism in colon cancer cells inhibition, staining was done using Hoechst 33342 (Thermofisher Scientific, MA, USA). WiDr and HCT-116 viable cells were seeded in an 8-well plate with a density of 1.0 x 104 cells/well and incubated at 37 °C in 5% CO2. The test concentration was similar to gene expression analysis. Following sterile PBS wash (3 times), 200 µL of 2 µM Hoechst stain was added, and they were incubated for one hour incubation at 37 ° C in dark condition. The dye was discarded and the cells were washed with PBS three times and allowed to dry. The observation was performed under a fluorescence microscope with a UV filter and the images were captured with Dino eyes pieces camera 1280×1024. The experiment was carried out in duplicate.
Statistical analysis
Data in the tables and figures were reported as mean ± SD. The result was analyzed using GraphPad Prism Version 8.30 (San Diego, USA). Statistical significance for cytotoxicity analysis was calculated using one-way or two-way ANOVA followed by the Sidak multiple comparison test. Significantly different values were determined when p<0.05. Gene expression and Hoechst staining result were presented in an average of the duplicate measurements.
Results
Cytotoxicity of Andaliman extracts
Andaliman extract exhibited inhibition activity in colon cancer cells in vitro indicated by the smaller number of cells were detected as the concentration of extracts was increased. Dox 3 μg.mL-1 suppressed the proliferation of colon cancer cells as expected. Interestingly, Dox caused a different response in the WiDr and HCT-116 cells (Table 2). Figure 1 showed the percentage of cell inhibition by Andaliman extract. Andaliman extract treatment exhibited inhibition in both HCT-116 and WiDr, with slightly different IC50 of 94.64 and 95.6 μg.mL-1, respectively. However, Andaliman extract showed lower inhibition in HCT-116 cell compared to the Dox treatment.
Table 2: Inhibition of WiDr and HCT-116 cells by Dox 3 and 1 μg.mL-1 as the control positive of cytotoxicity test
| Control + | % Inhibition | |
| WiDr | HCT-116 | |
| Dox 3 μg.mL-1 | 12.49 ± 2.13 | 77.91 ± 0.99 |
| Dox 1 μg.mL-1 | 1.38 ± 1.96 | 86.74 ± 2.09 |
Data were mean ± SD with n =3
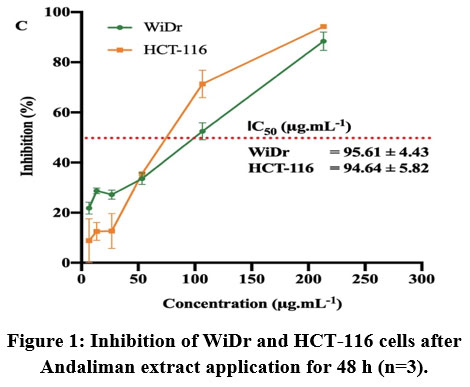 |
Figure 1: Inhibition of WiDr and HCT-116 cells after Andaliman extract application for 48 h (n=3). |
Mechanism of WiDr and HCT-116 cells inhibition
A study on pro-apoptosis (Bax) and anti-apoptosis (Bcl-2) markers was done using Real-Time PCR to investigate the mechanism of the colon cancer cells inhibition in vitro. In addition, Hoescht staining was used further to investigate the extent of this mechanism of action. WiDr and HCT-116 cells generally exhibited different responses toward Andaliman extract, which was consistent with the inhibition by Dox (Table 2).
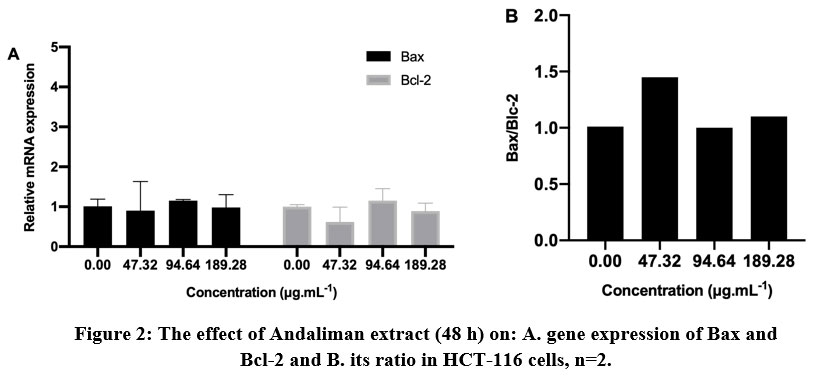 |
Figure 2: The effect of Andaliman extract (48 h) on: A. gene expression of Bax and Bcl-2 and B. its ratio in HCT-116 cells, n=2. |
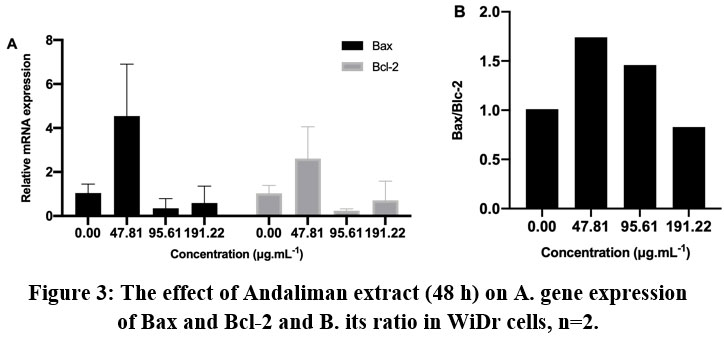 |
Figure 3: The effect of Andaliman extract (48 h) on A. gene expression of Bax and Bcl-2 and B. its ratio in WiDr cells, n=2. |
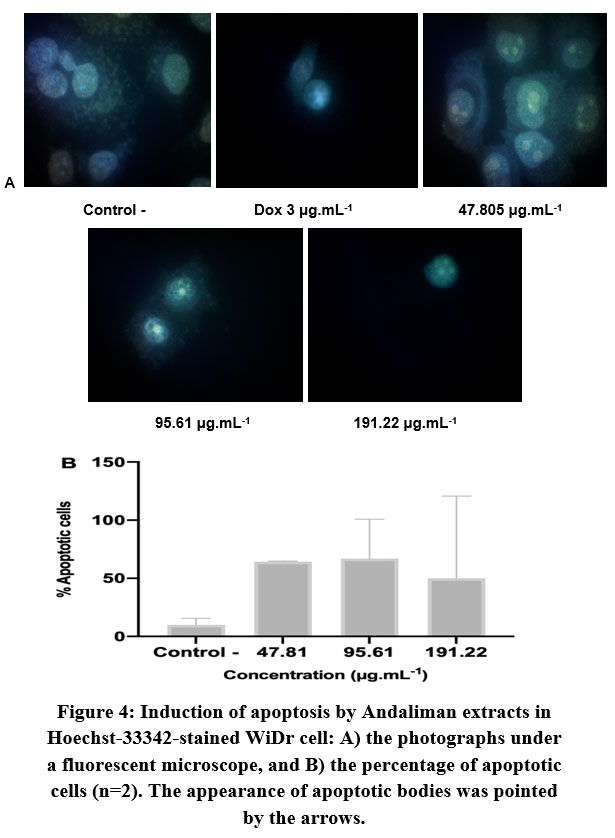 |
Figure 4: Induction of apoptosis by Andaliman extracts in Hoechst-33342-stained WiDr cell: A) the photographs under a fluorescent microscope, |
As seen in Figure 2A, the expression of both Bax and Bcl-2 were relatively low in HCT-116 cell. Bax/Bcl-2 coefficient was commonly used to evaluate the significant correlation of gene expression to the apoptotic status28. The ratio of Bax/Bcl-2 >1 showed apoptosis induction in the cells treated with a lower concentration of Andaliman extract (Figure 2B). To confirm this, we carried out Hoechst staining to mark the condensation or fragmentation of chromatin in the cells under a fluorescence microscope. Unfortunately, the apoptosis mechanism was not expressed in Hoechst-stained HCT-116 cells in any three treatments (data is not shown). Thus, the confirmation of the apoptosis mechanism from HCT-116 cells was inconclusive.
The expressions of Bax and Bcl-2 in WiDr cells were relatively higher than in HCT-116 cells (Figure 3). The significant expression of Bax and Bcl-2 in WiDr can be observed at the lowest concentration of extract. The application of higher concentration did not enhance the higher gene expression. However, the ratio of Bax/Bcl-2 showed that both treatments at concentrations of 47.81 μg.mL-1 and 95.61 μg.mL-1 induced apoptosis mechanism with a ratio value of 1.74 and 1.46, respectively, but not in higher concentration (191.22 μg.mL-1). Hoechst staining analysis (Figure 4) further supported the above observation.
Discussion
In this study, the cytotoxic potency of Andaliman extract toward human colon cancer cells was confirmed by the in vitro MTT method using HCT-116 and WiDr cell lines. These cells were isolated from different CRC patients with different mutation statuses of cancer-causing genes, such as the TP53 gene type, affecting their sensitivity toward different therapeutic agent29,30. From the IC50 value (Figure 1), Andaliman extracts inhibited both HCT-116 and WiDr cells effectively in the similar degree. Positive control (Dox) also exhibited a more effective inhibition in HCT-116 cell, showing this cell was more susceptible than WiDr cell (Table 2). Methanol, hexane, and ethyl acetate extract of Curcuma zedoaria with concentrations of 68.4, 41.1 and 21.0 μg.mL-1 could inhibit 50% of HCT-116 cells in vitro31, while Andaliman extract could perform a similar inhibition at a concentration of 94.64 μg.mL-1. Although Andaliman extract had a higher IC50, this study demonstrated that Andaliman effectively inhibited colon cancer in vitro.
Similar bioactivity was also observed in the essential oil extracted from Z. bungeanum leaves. It could inhibit the proliferation of SW620 cells with IC50 of 72.35 and 36.41 μM at 24 and 48 hours, respectively32. Nozaki et al. 33 found that Japanese pepper (Z. piperitum De Candolle) fruit extract could induce autophagic cell death (ACD) in the DLD-1 and Caco-2 cells at a concentration of 200 μg.mL-1. Interestingly, it did not give the same effect in WiDr cells. Inhibition toward Caco-2 colon adenocarcinoma cells was also shown by Z. armatum fruit, leaves, and bark with approximate IC50 of 200 μg.mL-1 for each type of methanolic extract. At a concentration of 200 μg.mL-1, the crude saponin from Z.armatum fruit extract could inhibit 75.88% of Caco-2 cells34.
These studies supported the finding above that Andaliman also posed bioactivity on the gastrointestinal tract, in this case, inhibition of colon cancer cells in vitro. No previous studies have reported Z. acanthopodium fruit bioactivity toward colorectal cells or its underlying cell death mechanism. Up to now, our study was the first to report the bioactivity against colorectal cancer cells.
Sanshool (250 μg.mL-1) isolated from Z. bungeanum, as one of the trigeminal sensation compounds known in Zanthoxylum species1, also showed inhibition toward hepatocarcinoma HepG2 cells with the inhibitory effect of 73.01% (48 hours)35. Sanshool compounds were also known as the major secondary amides found in Andaliman36. However, none study has proved the bioactivity of these compounds on colorectal cancer. Hydroxy-alpha-sanshool (HAS) has been used as one of the main constituents in Daikenchuto, a Japanese traditional medicine, that had a vasodilatory effect in 2,4,6-trinitrobenzenesulfonic acid-induced Crohn’s disease rat by enhancing the release of adrenomedullin3. It also gave a strong contraction effect in the rat proximal colon indicating its therapeutic function for colonic health4. Conversely, HAS and hydroxy-beta-sanshool (HBS) did not show any anticancer activity on DLD-1 colon cancer cells33. Although we did not examine the bioactive compound in this context; we speculated that the sanshool compounds might have contributed to the bioactivity of Andaliman fruit extract.
There were several pathways of cell death. One of the critical executions is programmed cell death, known as apoptosis, which can be induced intrinsically and extrinsically. In intrinsic regulation, the caspases group is the executioner. Meanwhile Bcl (B cell lymphoma)-2 family is the arbiter for cell death. The Bcl-2 proteins family is the backbone of the apoptosis mechanism and can regulate apoptosis negatively or positively. The opposing apoptosis proteins own 3-4 Bcl-2 Homology (BH) regions located at the outer mitochondrial membrane, endoplasmic reticulum and nuclear membrane37. The imbalance of pro-apoptotic and anti-apoptotic protein would induce tumorigenesis by inhibiting the apoptosis in cancer cells38. In this study, relative mRNA expression of Bax was markedly higher than Bcl-2, indicating that apoptosis through the extrinsic pathway was underlying the colon cancer cells’ inhibition (Figure 5). At least, in this case, Andaliman extract interfered and caused the cells’ death via mitochondrial disruption. A study found that Bax was more upregulated in the isolated tissue from patients in the Dukes’ C and D colon cancer stage than Dukes’ A and B, which was more likely shown in WiDr and HCT-116 cells39. A similar mechanism was also reported in the inhibition of SW620 cancer cells32, HepG2 cancer cells35 by Z. bungeanum, and LNCaP cancer cells by Zanthoxyli Fructus40. The downregulation of Bcl-2 protein and upregulation of Bax by Z. bungeanum essential oil was also reported in HaCaT cells41. Contrarily, the underlying mechanism of cell death induced by Z. piperitum fruit extract was autophagy, which was shown by the increased level of LF3-II protein and cytoplasmic vacuolization in DLD-1 cells33.
Apoptotic cells were usually marked by cell morphological changes, condensed chromatin, fragmented nuclei, and cell reduction42, which can be seen using Hoechst staining. Hoechst 33342 is a blue fluorescing dye that can stain chromatin DNA. This dye can be used to observe the features of apoptotic cells under a fluorescence microscope or phase contrast inverted microscope. By this analysis, we could confirm the apoptosis mechanism in WiDr cells by Andaliman extract. The concentration of 95.61 μg.mL-1 had the highest percentage of the apoptotic cell, yet it was close to the number of apoptotic cells at a concentration of 47.81 μg.mL-1, which was in line with the result of apoptosis gene expression analysis (Figure 3). Although reviewed as one of the most popular and excellent fluorescent chemical compounds used to detect the apoptotic cells, Hoechst staining has some limitations. Visualizing in the UV region may be interfered by tissue autofluorescence and phototoxicity of the UV light, peculiarly when there is only a small amount of non-nuclear DNA detected in the sample43. This might appear as the issue in the HCT-116 cells observation in this research.
The expression of Bax and Bcl-2 after Andaliman extract applications were not in a dose-dependent manner in both cells (Figure 2 and Figure 3). An increasing dose of Andaliman extract reduced the expression of the apoptotic gene marker. Similar trend was also shown by Trolox application which had dual behavior as an antioxidant and prooxidant agents on the model HeLa cells over wide concentrations. It had induced prooxidant activity at concentration above 15 μM which was accompanied by the decrease of cell viability due to apoptosis44. Similar dual roles was also observed in the other cancer therapeutic studies. In the specific concentrations, reactive oxygen species (ROS) can induce cancer cell death by several pathways, for instance the activation of ASK1/JNK and ASK1/p38 signaling pathways45 and inactivation of glyceraldehyde 3-phosphate dehydrogenase (GAPDH) in colon cancer with KRAS or BRAS mutations in vitro46. Therefore, we speculate that this trend was potentially caused by the alteration of intracellular ROS due to different concentrations of Andaliman extracts on the observed colon cancer cells.
Interestingly, the expression of apoptotic gene markers in the WiDr cell tended to be higher than in the HCT-116 cell. Based on the cell origin, WiDr and HCT-116 were distinguished by the colon cancer staging. HCT-116 was isolated from colorectal carcinoma patients at Dukes’ stage D; meanwhile, WiDr was from colorectal adenocarcinoma patient at Dukes’ stage C. The Bax/Bcl-2 coefficient tended to increase with tumor progression39, i.e., with more advancing in growth and metastases, which was not precisely shown in this research. Nevertheless, the difference was tight between Dukes’ C and D, as shown in the previous study39. Both cells also shared a different molecular pathway and mutation status29. These features might be the reason for the differences in the inhibition trend and the gene expression between the two cell lines. WiDr, LS-174, and HT-29 cell was reported to overexpress COX-2, whereas HCT-116 did not express COX-247. Palozza et al. observed that not expressing COX-2 cells (HCT-116) was insensitive to the growth inhibitory and pro-apoptotic effect of ß-carotene treatment47. A similar phenomenon was suspected as the underlying mechanism of higher expression of pro-apoptotic gene marker in WiDr than HCT-116 cells in this study.
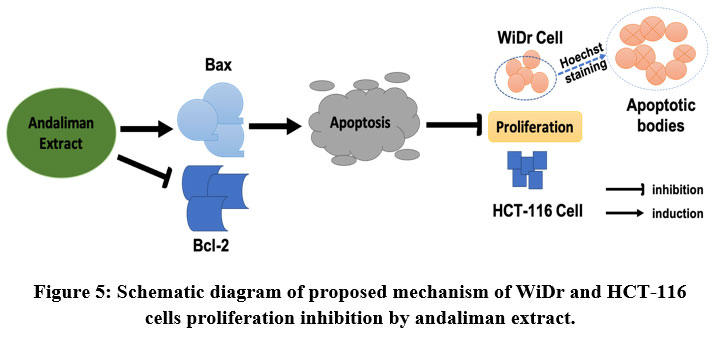 |
Figure 5: Schematic diagram of proposed mechanism of WiDr and HCT-116 cells proliferation inhibition by andaliman extract. |
There are several limitations to this study that should be considered. Only Bcl-2 and Bax gene markers were used to assess the underlying mechanism of colon cancer cells inhibition by Andaliman extract. Other apoptotic markers, such as PARP cleavage and several types of caspases, can also be used to validate the apoptosis mechanism at the protein level in this experiment. Besides, cell cycle analysis and cell cycle regulatory protein expression can be performed to support the result. Furthermore, an animal study has not been conducted yet. In vivo studies can give more comprehensive analysis of Andaliman extract’s bioactivity to prevent colon cancer disease.
Conclusion
Andaliman ethanolic extract possessed bioactivity toward colorectal cancer cells inhibition in vitro, indicating its potency to be developed as a functional food ingredient for colon cancer disease prevention. The suggested mechanisms of action behind this inhibition were apoptosis and not in a dose-dependent manner. Moreover, the trend was different between the studied cells. For future research, it would be interesting to see the compounds inside the extract responsible for this bioactivity. Cell cycle analysis, additional apoptotic markers and cell cycle regulatory protein expression, as well as in vivo experiment are suggested to be done as a validation approach of our finding here.
Acknowledgement
This research was conducted at IPB University. The authors would like to thank the Primate Research Center IPB for the professional assistance with the colon cancer cells in vitro studies.
Conflict of Interest
The authors declare no conflict of interest.
Funding Sources
This research was financially supported by Ministry of Research, Technology, and Higher Education of Indonesia (Kemenristekdikti) under PMDSU (Pendidikan Magister Menuju Doktor untuk Sarjana Unggul) Scheme (Grant No. 025/E3/2017).
Reference
- Wijaya C.H., Wijaya Napitupulu F.I., Karnady V., Indariani S. A Review of the Bioactivity and Flavor Properties of The Exotic Spice “Andaliman” (Zanthoxylum acanthopodium DC.). Food Rev. Int. 2019; 35(1):1-19. doi:10.1080/87559129.2018.1438470
CrossRef - Napitupulu B., Simatupang S., Sinaga M. Potency of Andaliman as Traditional Food Additive of Batak Ethnic North Sumatera. Paper presented at: Seminar Nasional Peningkatan Daya Saing Pangan Tradisional; 2004; BB Pascapanen Bogor, Indonesia.
- Kono T., Omiya Y., Hira Y., Kaneko A., Chiba S., Suzuki T., Noguchi M., Watanabe T. Daikenchuto (TU-100) Ameliorates Colon Microvascular Dysfunction via Endogenous Adrenomedullin in Crohn’s Disease Rat Model. J. Gastroenterol. 2011; 46(10):1187-1196. doi:10.1007/s00535-011-0438-2.
CrossRef - Kubota K., Ohtake N., Ohbuchi K., Mase A., Imamura S., Sudo Y., Miyano K., Yamamoto M., Kono T., Uezono Y. Hydroxy-α Sanshool Induces Colonic Motor Activity in Rat Proximal Colon: A Possible Involvement of KCNK9. Am. J. Physiol. Gastrointest. Liver Physiol. 2015; 308(7):G579-G590. doi:10.1152/ajpgi.00114.2014
CrossRef - Zhang Z., Liu J., Shen P., Cao Y., Lu X., Gao X., Fu Y., Liu B., Zhang N. Zanthoxylum bungeanum Pericarp Extract Prevents Dextran Sulfate Sodium-Induced Experimental Colitis in Mice via the Regulation of TLR4 and TLR4-related Signaling Pathways. Int. Immunopharmacol. 2016; 41:127-135. doi:10.1016/j.intimp.2016.10.021
CrossRef - Zhang Z., Shen P., Liu J., Z., Shen P., Liu J., Gu C., Lu X., Li Y., Cao Y., Liu B., Fu Y., Zhang N. In Vivo Study of the Efficacy of the Essential Oil of Zanthoxylum bungeanum Pericarp in Dextran Sulfate Sodium-Induced Murine Experimental Colitis. J. Agr. Food Chem.. 2017; 65(16):3311-3319. doi:10.1021/acs.jafc.7b01323
CrossRef - Gultom S. Flavonoid of Zanthoxylum achanthopodium Fruit as an Antioxidant and α-Glucosidase Inhibitors. 2011. MS Thesis, Institut Pertanian Bogor, Bogor, Indonesia.
- Suryanto E., Raharjo S., Sastrohamidjojo H., Tranggono. Anti-photooxidative Effect Of Andaliman (Zanthoxylum acanthopodium DC.) Extract on Linolenic Acid. Agritech. 2005; 24(4):173-179.
- Yanti, Pramudito T.E., Nuriasari N., Juliana K. Lemon Pepper Fruit Extract (Zanthoxylum acanthopodium DC.) Suppresses the Expression of Inflammatory Mediators in Lipopolysaccharide-Induced Macrophages In Vitro. Am. J. Biochem. Biotech. 2011; 7(4):190-195. doi:10.3844/ajbbsp.2011.190.195
CrossRef - Anggraeni R., Hadisahputra S,. Silalahi J., Satria D. Combinational Effects of Ethylacetate Extract of Zanthoxylum acanthopodium DC. with Doxorubicin on T47D Breast Cancer Cells. Int. J. PharmaTech Res. 2015; 6(7):2032-2035.
- Kristanty R.E., Suriawati J. The Indonesian Zanthoxylum acanthopodium DC.: Chemical and Biological Values. Int. J. PharmTech Res. 2015; 8(6):313-321.
- Karnady V. Comparison on In Vitro Digestion Effect of Antioxidant and Antihyperglycemic Activity from Andaliman (Zanthoxylum acanthopodium DC.) And Japanese Pepper (Zanthoxylum piperitum DC .) Crude Extract. 2015. MS Thesis, Institut Pertanian Bogor, Bogor, Indonesia.
- Hanum I., Laila L. Physical Evaluation of Anti-aging and Anti-acne Andaliman (Zanthoxylum acanthopodium DC.) Ethanolic Extract Peel Off Gel Mask. Der Pharma Chemica. 2016; 8(23):6-10.
- International Association of Cancer Research. The Global Burden of Cancer. https://gco.iarc.fr/today/online-analysis-table. Published 2018. Accessed November 17, 2020.
- International Agency for Research on Cancer. The Global Cancer Observatory: Cancer Incident in Indonesia. https://gco.iarc.fr/today/data/factsheets/ populations /360-indonesia-factsheets.pdf. Published March 2021. Accessed October 30, 2021.
- Tariq K., Ghias K. Colorectal Cancer Carcinogenesis: A Review of Mechanisms. Cancer Biol. Med. 2016; 13(1):120-135. doi:10.28092/j.issn.2095-3941.2015.0103
CrossRef - Szarynska M., Olejniczak A., Wierzbicki P., Kobiela J., Laski D., Sledzinski Z., Adrych K., Guzek M., Kmiec Z. FasR and FasL in Colorectal Cancer. Int. J. Oncol. 2017; 51(3):975-986. doi:10.3892/ijo.2017.4083
CrossRef - Mohammad N., Malvi P., Meena A.S., Singh S.V., Chaube B., Vannuruswamy G., Kulkarni M.J., Bhat M.K. Cholesterol Depletion by Methyl-β-Cyclodextrin Augments Tamoxifen Induced Cell Death by Enhancing Its Uptake in Melanoma. Mol. Cancer. 2014; 13(1):1-13. doi:10.1186/1476-4598-13-204
CrossRef - Mohammad N., Singh S.V., Malvi P., Chaube B., Athavale D., Vanuopadath M., Nair S.S., Nair B., Bhat M.J. Strategy to Enhance Efficacy of Doxorubicin in Solid Tumor Cells by Methyl-β-Cyclodextrin: Involvement of p53 and Fas Receptor Ligand Complex. Sci. Rep. 2015; 5 (11853):1-13. doi:10.1038/srep11853
- Hawk E.T., Levin B. Colorectal Cancer Prevention. J. Clin. Oncol. 2005; 23(2):378-391. doi:10.1200/JCO.2005.08.097
CrossRef - Rao S., Chinkwo K., Santhakumar A., Johnson S., Blanchard C. Apoptosis Induction Pathway in Human Colorectal Cancer Cell Line SW480 Exposed To Cereal Phenolic Extracts. Molecules. 2019; 24(13). doi:10.3390/molecules24132465
CrossRef - Jaksevicius A., Carew M., Mistry C., Modjtahedi H., Opara E.I. Inhibitory Effects Of Culinary Herbs And Spices on the Growth of HCA-7 Colorectal Cancer Cells and Their COX-2 Expression. Nutrients. 2017; 9(1051). doi:10.3390/nu9101051
CrossRef - Kirana C., Record I.R., McIntosh G.H., Jones G.P. Screening for Antitumor Activity of 11 Species of Indonesian Zingiberaceae Using Human MCF-7 and HT-29 Cancer Cells. Pharma. Bio. 2003; 41(4):271-276. doi:10.1076/phbi.41.4.271.15673
CrossRef - Napitupulu F.I.R, Wijaya C.H., Prangdimurti E., Akyla C., Yakhin L.A., Indriyani S. Comparison of Several Processing Methods in Preserving the Flavor Properties of Andaliman (Zanthoxylum acanthopodium DC.) Fruit. J. Eng. Technol. Sci. 2020; 52(3):1-14. doi: 10.5614/j.eng.technol.sci.2020.52.3.7
CrossRef - Livak K.J., Schmittgen T.D. Analysis of relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods. 2001; 25(4):402-408. doi:10.1006/meth.2001.1262
CrossRef - Borhani N., Manoochehri M., Saleh Gargari S., Ghaffari Novin M., Mansouri A., Omrani M.D. Decreased Expression of Proapoptotic Genes Caspase-8- and BCL2-Associated Agonist of Cell Death (BAD) in ovarian Cancer. Clin. Ovarian Gynecol. Cancer. 2014; 7(1-2):18-23. doi:10.1016/j.cogc.2014.12.004
CrossRef - Yulianto W., Andarwulan N., Giriwono P.E., Pamungkas J. HPLC-based Metabolomics to Identify Cytotoxic Compounds from Plectranthus amboinicus (Lour.) Spreng Against Human Breast Cancer MCF-7 Cells. J.Chromatogr. B: Ana. Tech. Biomed. Life Sci. 2016; 1039:28-34. doi:10.1016/j.jchromb.2016.10.024
CrossRef - Khodapasand E., Jafarzadeh N., Farrokhi F., Kamalidehghan B., Houshmand M. Is Bax/Bcl-2 Ratio Considered as a Prognostic Marker with Age and Tumor Location In Colorectal Cancer? Iranian Biomed. J. 2015; 19(2):69-75. doi:10.6091/ibj.1366.2015
- Ahmed D., Eide P.W., Eilertsen I.A., Danielsen S.A., Eknæs M., Hektoen M., Lind G.E., Lothe R.A. Epigenetic and Genetic Features of 24 Colon Cancer Cell Lines. Oncogenesis. 2013; 2(e71):18. doi:10.1038/oncsis.2013.35
CrossRef - Giovannetti E., Backus H.H.J., Wouters D., Ferreira C.G., van Houten V.M.M, R Brakenhoff H., Poupon M-F., Azzarello A., Pinedo H.M., Peters G.J. Changes in the status of p53 affect drug sensitivity to thymidylate synthase (TS) inhibitors by altering TS levels. British J. Cancer. 2007; 96(5):769-775. doi:10.1038/sj.bjc.6603639
CrossRef - Syed Abdul Rahman S.N., Abdul Wahab N., Abd Malek S.N. In vitro Morphological Assessment of Apoptosis Induced by Antiproliferative Constituents from the Rhizomes of Curcuma zedoaria. Evid.-based Compl. Alt. Med. 2013; 2013: 1-14 doi:10.1155/2013/257108
CrossRef - Zhang Y., Dong H., Zhang J., Zhang L. Inhibitory effect of hyperoside isolated from Zanthoxylum bungeanum leaves on SW620 human colorectal cancer cells via induction of the p53 signaling pathway and apoptosis. Mol. Med. Rep.. 2017;16(2):1125-1132. doi:10.3892/mmr.2017.6710
CrossRef - Nozaki R., Kono T., Bochimoto H., Watanabe T., Oketani K., Sakamaki Y., Okubo N., Nakagawa K., Takeda H. Zanthoxylum fruit extract from Japanese pepper promotes autophagic cell death in cancer cells. Oncotarget. 2016; 7(43):70437-70446. doi:10.18632/oncotarget.11926
CrossRef - Alam F., Najum us Saqib Q., Waheed A. Cytotoxic Activity Of Extracts And Crude Saponins from Zanthoxylum armatum DC. against Human Breast (MCF-7, MDA-MB-468) and Colorectal (Caco-2) Cancer Cell Lines. BMC Com. Alt. Med. 2017; 17(1):1-9. doi:10.1186/s12906-017-1882-1
CrossRef - You Y., Zhou M., Lu H., Shirima G.G., Cheng Y., Liu X. Sanshool from Zanthoxylum L. Induces Apoptosis In Human Hepatocarcinoma HepG2 Cells. Food Sci. Biotech. 2015; 24(6):2169-2175. doi:10.1007/s10068-015-0289-3
CrossRef - Suharta S., Hunaefi D., Wijaya C.H. Changes in Volatiles and Aroma Profile of Andaliman (Zanthoxylum acanthopodium DC.) upon Various Drying Techniques. Food Chem. 2021; 365(130483):1-10. doi:10.1016/j.foodchem.2021.130483
CrossRef - Pellegrini M., Strasser A.. Caspases, Bcl-2 Family Proteins and Other Components of the Death Machinery: Their Role in the Regulation of the Immune Response. In: Madame Curie Bioscience Database. Austin (TX): Landes Bioscience; 2013. https://www.ncbi.nlm.nih.gov/books/NBK6432/. Accessed February 11, 2020.
- Bostan M., Mihaila M., Hotnog C., Bleotu C., Anton G., Roman V., Brasoveanu L.I. Colorectal Cancer: From Pathogenesis to Treatment. In: Rodrigo L, ed. Intechopen.; 2016. doi:http://dx.doi.org/10.5772/63382 87
CrossRef - Paul-Samojedny M., Kokocińska D., Samojedny A., Mazurek U., Partyka R., Lorenz Z., Wilczok T. Expression of Cell Survival/Death Genes: Bcl-2 and Bax at the Rate of Colon Cancer Prognosis. Biochimica et Biophysica Acta 2005; 1741:25-29. doi:10.1016/j.bbadis.2004.11.021
CrossRef - Yang Y., Ikezoe T., Takeuchi T., Adachi Y., Ohtsuki Y., Koeffler H.P., Taguchi H. Zanthoxyli Fructus Induces Growth Arrest and Apoptosis of LNCaP Human Prostate Cancer Cells In Vitro and In Vivo in Association with Blockade of the AKT and AR Signal Pathways. Oncol. Reports. 2006; 15(6):1581-1590. doi:10.3892/or.15.6.1581
CrossRef - Li K., Zhou R., Jia W.W., Li Z., Li J., Zhang P., Xiao T. Zanthoxylum bungeanum Essential Oil Induces Apoptosis of HaCaT Human Keratinocytes. J. Ethnopharmaco. 2016; 186:351-361. doi:10.1016/j.jep.2016.03.054
CrossRef - Crowley L.C., Marfell B.J., Waterhouse N.J. Analyzing Cell Death by Nuclear Staining with Hoechst 33342. Cold Spring Harbor Protocols. 2016; 2016(9):778-781. doi:10.1101/pdb.prot087205
CrossRef - Bucevičius J., Lukinavičius G., Gerasimaite R. The Use of Hoechst Dyes for DNA staining And Beyond. Chemosensors. 2018; 6(2). doi:10.3390/chemosensors6020018
CrossRef - Giordano M.E., Caricato R., Lionetto M.G. Concentration Dependence of the Antioxidant and Prooxidant Activity of Trolox in HeLa Cells: Involvement in the Induction of Apoptotic Volume Decrease. Antioxidants. 2020; 9(11):1-12. 10.3390/antiox9111058.
CrossRef - Reczek C.R., Chandel N.S. The Two Faces of Reactive Oxygen Species in Cancer. Annu. Rev. Cancer Biol. 2017; 1:79-98. doi:https://doi.org/10.1146/annurev-cancerbio-041916-065808
CrossRef - Yun J., Mullarky E., Lu C., et al. Vitamin C Selectively Kills KRAS and BRAF Mutant Colorectal Cancer Cells by Targeting GAPDH. Sci. 2015; 350(6266):1391-1396. doi:doi:10.1126/science.aaa5004.
CrossRef - Palozza P., Serini S., Maggiano N., Tringali G., Navarra P., Ranelletti F.O, Calviello G. β-Carotene Downregulates the Steady-State and Heregulin-α–Induced COX-2 Pathways in Colon Cancer Cells. Nut. Cancer. 2005; 135(1):129-136. doi:10.1093/jn/135.1.129
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








