Introduction
Obesity is a well-defined epidemic in Westernized cultures and increasing prevalence is being seen in developing countries too. With obesity comes a variety of adverse health outcomes, such as high blood pressure, insulin resistance, and type 2 diabetes. Many lines of evidence have shown that chronic activation of proinflammatory pathways within insulin target cells can lead to obesity-related insulin resistance. Consistent with this, elevated levels of the proinflammatory cytokines TNFα, IL-6, and C-reactive protein (CRP) have been shown in individuals with insulin resistance and diabetes.1Insulin resistance is defined as an inadequate response by insulin target tissues, such as skeletal muscle, liver, and adipose tissue,to the physiologic effects of circulating insulin. The hallmarks of impaired insulin sensitivity in these three tissues are decreased insulin-stimulated glucose uptake into skeletal muscle, impaired insulin-mediated inhibition of hepatic glucose production in liver, and a reduced ability of insulin to inhibit lipolysis in adipose tissue.In fact, insulin resistance is a major predictor for the development of various metabolic sequelae, including type 2 diabetes and is a defining feature of syndrome X, which is also known as the metabolic syndrome. This syndrome encompasses a constellation of conditions, including insulin resistance, dyslipidemia, hypertension, and obesity, and is often accompanied by hyperinsulinemia, sleep apnea, and other disorders.2 MetS increases the risk developing CVD by 2-fold. Obesity (BMI > 30 kg/m2), which presents with dyslipidemia and elevated cholesterol levels, plays a major role in the development of MetS, which increases the risk of type II diabetes. Because of the exploding obesity epidemic, research efforts have escalated to better understand all aspects of the pathophysiology, including how obesity affects lipid and lipoprotein metabolism.
Numerous factors affect variability in lipid response to diet.Greater cholesterol synthesis and lower cholesterol absorption can arise as a consequence of insulin resistance, often causing a diminished plasma lipid response to diet.3 Dietary interventions are effective for improving the lipid and lipoprotein profile. Replacing SFA with MUFA, PUFA, and/or carbohydrate (CHO) reduces LDL cholesterol (LDL-C).4,5 A mechanism by which replacing SFA with PUFA lowers LDL-C is via an increase in LDL receptor(LDLR)-mediated uptake of LDL-C from circulation .LDLR-mediated uptake, however, is impaired by obesity.6 Consequently, obese individuals are less responsive to dietary interventions aimed at improving the lipid/lipoprotein profile. A greater understanding of the factors that diminish lipid uptake in obese individuals likely will increase understanding of why they have a blunted lipid response to dietary interventions.
The purpose of this review is to summarize the evidence and discuss the possible mechanisms that contribute to the blunted lipid response to dietary change that is associated with obesity. This review will discuss the effects of dietary SFA and cholesterol on changes in plasma lipids and lipoproteins and the effect of adiposity on these responses. However endocrine disregulation is beyond the scope of this review
Obesity Causes A Blunted Lipid Response to SFA and Cholesterol in the Diet
A number of clinical studies have shown an inverse relation between BMI and lipid response. The lipid and lipoprotein response is greater in lean individuals compared to obese. Bronsgeest-Schoute et al., included participants who typically consumed at leat 1 egg/d.They were not allowed to consume any egg products for three weeks. The experimental diet consisted of 264mg/d cholesterol and their daily diet consisted of 742mg/d. A small but significant decrease in total cholesterol (TC) was reported (−0.16 ± 0.40 mmol/L; P < 0.05) with an inverse correlation (r = −0.321; P < 0.05) between BMI and the reduction in TC. When participants were classified on the basis of BMI, only those who were not obese had a reduction in TC (−0.23 ± 0.43 mmol/L; P < 0.01.7
A study done by Mukuddem- Petersen et al. reported no lipid/lipoprotein effects of a weight maintenance diet that contained either walnuts or cashews (20% of energy came from nuts) in obese individuals. The study showed that the benefit of nut consumption was not seen in obese individuals having metabolic syndrome.8
Lefevre et al did a randomized, double-blind, 3-period crossover controlled feeding design to examine the effects on plasma lipids of 3 diets that differed in total fat: the average American diet (AAD) [designed to contain 38% fat and 14% saturated fatty acids (SFAs)], the Step I diet (30% fat with 9% SFAs), and the Step II diet (25% fat with 6% SFAs). The diets were fed for 6 wk each to 86 free-living, healthy men aged 22–64 y at levels designed to maintain weight. This study showed that persons who are insulin resistant respond less favorably to Step II diets than do those who are insulin sensitive. The data suggested that even in the absence of overt metabolic syndrome, weight reduction may be required to fully derive the benefit of dietary changes on CVD risk.9
A study done by Jansen et al, aimed at establishing whether being overweight is a factor whoch determines individuals response to dietary lipid changes. Forty-one non-obese healthy men were divided into two groups according to body mass index as follows: controls, <25 kg/m2.
overweight, >25 kg/m2 but <30 kg/m2. After consuming a saturated fat–rich diet (SAT diet: 38% fat, 20% saturated) for 4 wk, subjects were switched to a low fat diet [National Cholesterol Education Program (NCEP)-I diet: 28% fat, 10% saturated] for 4 wk and then to a monounsaturated fat–rich diet (MUFA diet: 38% fat, 22% monounsaturated) for 4 week. Plasma cholesterol concentrations changed to a lesser extent, and triglyceride concentration to a greater extent, in overweight but non-obese young men than in those of normal weight in response to changes in dietary fat composition. The data suggest that in the diet treatment of obese hyperlipemic subjects, it is more important for them to lose weight than to change the fat composition of their diets.10
From the above studies it is seen that body weight is an important predictor of response to changes in diet quality of individuals. The plasma cholesterol lowering effect of low SFA and cholesterol diet was more in lean individuals as compared to obese individuals
Adipose Tissue Induced Inflammation
Excess adipose tissue results in inflammation that leads to insulin resistance. The increase in adipocyte size and ensuing expansion of adipose tissue mass increases FFA release into the circulation and decreases oxygen delivery to the cells.2 This leads to an increase in cellular stress, adipocyte death, and expression of inflammatory genes, enhancing the activation of the proinflammatory c-Jun N-terminal kinase 1 (JNK1) and inhibitor of kappaB kinase (IKK)/NF-κB pathways.11 Macrophages accumulate in the adipose tissue and remodel the tissue.2 Additional proinflammatory cytokines (TNFα and IL-6) and chemokines are released, which can initiate the JNK1 and IKK/NF-κB pathways in nearby adipocytes, causing further macrophage recruitmentto local sites of injury, or circulate to the liver and initiate a similar process. Adipocytes also secrete a variety of adipokines, many of which affect insulin sensitivity. For instance, leptin and adiponectin have been shown to promote insulin sensitivity, whereas resistin and retinol-binding protein 4 interfere with insulin action and diminish insulin sensitivity. Eventually, an inflammatory environment in insulin target cells, specifically adipocytes and hepatocytes, causes localized insulin resistance due to stimulation of adipocyte lipolysis and complications associated with the inflammatory response, including reduced adiponectin, increased resistin, and increased hepatic glucose production.2
The proinflammatory cytokine TNFα can contribute independently to insulin resistance by reducing insulin receptor expression, insulin receptor substrate and GLUT4 gene expression, adiponectin, hormone sensitive lipase, and insulin-mediated glucose uptake.12,13 CRP is synthesized and secreted by the liver in response to proinflammatory cytokines, specifically IL-6.TNFα, IL-6, and CRP typically are elevated in insulin-resistant states.14 Human serum paraoxonase 1 (PON1) is an enzyme with esterase activity, and is physically bound to high-density lipoproteins (HDL). It plays a key role in the action of HDL toward protection of lipoprotein and biological membrane against oxidative damage In obese adults, diminished levels of PON1 activity is correlated with low levels of HDL cholesterol.15 In an obese individual, lipid also can accumulate in the muscle and liver independent of adipocyte lipolysis, initiating a proinflammatory state and the development of insulin resistance. Inflammation has been demonstrated to impair reverse cholesterol transport (RCT) at various steps in the pathway, conserving cholesterol stores in the body and preventing cholesterol flux through liver to bile and feces.16 The inhibition of RCT likely contributes to insulin resistance and MetS, negatively altering the lipid profile and potentially accelerating the development of CVD.
The key point is that excess adipose tissue and nutrient intake causes an increase in inflammation, which leads to the development of insulin resistance and the ensuing decrease in lipid response to changes in dietary SFA and cholesterol.
Obesity, Tissue Inflammation, and Insulin Resistance
Nutrient excess, weight gain, and ensuing obesity result in expansion of adipose tissue mass and adipocyte size. With this expansion, total free fatty acid release into the circulation is increased and oxygen delivery to the adipocyte is decreased. The combination of microhypoxia and nutrient excess leads to induction of Hypoxia inducible factor-1 HIF-1 and the downstream target genes as well as endoplasmic reticulum (ER) stress within the adipocyte. This can lead to the eventual death of the adipocyte as well as a characteristic inflammatory response. The inflammatory response includes increased production and release of proinflammatory cytokines/chemokines and the recruitment of bone marrow–derived macrophages. These macrophages are of the M1 activation/polarization state and are highly inflammatory in nature. Once recruited, these macrophages release proinflammatory cytokines, which work in a paracrine manner to activate the intracellular proinflammatory pathways (e.g., JNK and IKK) in neighboring cells and possibly through endocrine mechanisms in distal tissues. In a feed-forward cycle, activation of macrophages promotes the recruitment and infiltration of additional macrophages into adipose tissue. This results in cell autonomous insulin resistance in adipocytes and liver, exacerbation of the inflammatory state, and systemic insulin resistance. With obesity, there is also increased fat accumulation within skeletal muscle, and these intermuscular fat depots becomes infiltrated with proinflammatory macrophages, which may cause paracrine-like insulin resistance in skeletal muscle. In parallel with these inflammation-related changes, alterations in fatty acid metabolism can lead to the accumulation of fatty acid intermediates with the liver and skeletal muscle, which can cause insulin resistance. (Figure-1).2
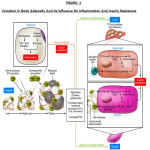 |
Figure 1: Variation In Body Adiposity And Its Influence On Inflammation And Insulin Resistance Click here to View figure |
Ref: Insulin sensitivity: modulation by nutrients and inflammation
Simon Schenk, Maziyar Saberi, Jerrold M. Olefsky Published in Volume 118, Issue 9 J Clin Invest. 2008; 118(9):2992–3002 doi:10.1172/JCI3426
Insulin Resistance Promotes Lipid Synthesis and Excretion
Insulin resistance or elevated insulin concentrations directly lead to metabolic changes in the hepatocyte that result in diminished responsiveness of the hepatic LDL-cholesterol receptor to changes in dietary fatty acids. This occurs through a mechanism whereby elevated insulin concentrations directly increase hepatic cholesterol synthesis through activation of the liver X receptor. Secondary to the elevated endogenous production of cholesterol, hepatic LDL-cholesterol receptors would be expected to be down-regulated and therefore refractory to additional reductions in LDL cholesterol that are typically associated with dietary changes.17 LXRs have been proposed to act as sterol sensors that function to help the organism adapt so it can cope with the effects of high free cholesterol levels in blood.18 The activity of lipoprotein lipase (LPL) is an important first step in plasma triglyceride clearance and FFA delivery to the adipocyte, particularly in the postprandial state. Insulin and glucose have been shown to stimulate adipose tissue LPL activity and to reduce LPL activity in muscle, implying a preferential postprandial partitioning of lipoprotein- derived fatty acids toward adipose tissue and away from muscle. In obesity and type 2 diabetes, insulin activation of LPL in adipose tissue is delayed, and LPL activity in skeletal muscle is increased instead of decreased by hyperinsulinemia. The importance of LPL in tissue FFA uptake has recently been demonstrated by experiments in which either muscle-specific or liver-specific overexpression in mice induces marked tissue lipid accumulation.19 Although LPL may be viewed as a first step leading to the uptake of FFA by adipose tissue, it is clear that the deposition of FFA is also regulated downstream of LPL. Thus in the presence of insulin resistance, LDLR activity is blunted and LDL binding declines, resulting in impaired receptor-mediated LDL-C removal and decreased chylomicron remnant clearance.17 The decrease in LDL-C uptake leads to an increase in endogenous cholesterol production, perhaps by the stimulation of LXRα. Hyperinsulinemia has been shown to stimulate LXRα which is known to regulate lipogenesis and cholesterol excretion. However, it is possible that decreased cholesterol absorption is secondary to increased cholesterol synthesis. Both states are related to insulin resistance; therefore, concurrent changes in both cholesterol absorption and synthesis make it difficult to determine which state is affected primarily by insulin resistance. Greater cholesterol production likely would lead to a further decline in LDLR activity and, consequently, a resistance to reductions in LDL-C that are associated with dietary fat and cholesterol modifications.20
ATP-binding cassette (ABC) transporters,Abcg5 and Abcg8 are expressed predominantly in the liver and small intestine and are coordinately up-regulated at the transcriptional level by dietary cholesterol. The response of Abcg5 and Abcg8 to cholesterol requires the liver X receptor α (LXRα) .Disruption of insulin signaling and stimulation of the LXRα pathway increases the expression of intestinal, specifically ABCG5 and ABCG8.21 ABCG5 and ABCG8 regulate the secretion of cholesterol and sterols from intestinal enterocytes into the intestinal lumen and from hepatocytes into the biliary space.22 Therefore, upregulation of ABCG5 and ABCG8 promotes biliary cholesterol secretion and decreased cholesterol absorption, which leads to the increase in hepatic cholesterol synthesis.23 High-cholesterol and high-fat diets also have been shown to increase the mRNA of ABCG5 and ABCG8. Subtle defects in these proteins or in their regulation may underlie the variable responses of healthy individuals to high-cholesterol diet.24 In addition, single nucleotide polymorphisms in the ABCG5 and ABCG8 genes can alter cholesterol metabolism and various lipid responses.25 Thus, ABCG5 and ABCG8 are important factors to consider in regulating endogenous cholesterol homeostasis. Hyperinsulinemia enhances expression of the ABCG5 and ABCG8 genes, stimulating cholesterol excretion and decreasing cholesterol absorption. However, proinflammatory cytokines, IL-6 and TNFα, as well as insulin, have been shown to inhibit CYP7A1 gene transcription, thereby decreasing bile acid synthesis as an adaptive response to protect hepatocytes from injury.Evidence suggests that these cell-signaling pathways cross talk to regulate bile acid synthesis to maintain hepatic bile acid homeostasis.26
Insulin resistance initiated by adipose tissue-induced inflammation leads to increased cholesterol synthesis and decreased cholesterol absorption
Hypocaloric Diets and Lipid Response Lipid Response in Obese Individuals
The lipid and lipoprotein response is greater in obese individuals following weight loss.27-31 Following are a few selected studies which have reported that hypocaloric diet in addition to weight loss also have improved the lipid profile in obese and overweight individuals.
Table 1: Selected Researches on Effect of Hypocaloric Diets in Obese Individuals
| Article | Author and Year | N (M/F) | Method | Observation | Conclusion |
| The effects of a whole grain–enriched hypocaloric diet on cardiovascular disease risk factors in men and women with metabolic syndrome | Heather I Katcher et al ,2008,AJCN | 50(25/25 )Obese Adults | Obese adults with metabolic syndrome were randomly assigned to receive dietary advice either to avoid whole-grain foods or to obtain all of their grain servings from whole grains for 12 wk. All participants were given the same dietary advice in other respects for weight loss. . Energyneeds were calculated by using the Mifflin equation with an activity factor of 1.3, and subtracting 500 kcal to accountfor the calorie deficit needed to achieve weight loss | Body weight, waist circumference, and percentage body fat decreased significantly in both groups over the study period, but there was a significantly greater decrease in percentage body fat in the abdominal region in the whole-grain group than in the refined-grain group. C-reactive protein (CRP) decreased 38% in the whole-grain group independent of weight loss but was unchanged in the refined-grain group . Total, LDL, and HDL cholesterol decreased in both diet groups (P< 0.05). | Both hypocaloric diets were effective means of improving CVD risk factors with moderate weight loss. There were significantly (P< 0.05) greater decreases in CRP and percentage body fat in the abdominal region in participants consuming whole grains than in those consuming refined grains. |
| Article | Author and Year | N (M/F) | Method | Observation | Conclusion |
| The effect of hypocaloric diet enriched in legumes with or without L-arginine and selenium on anthropometric measures in central obese women | Mohammad Alizadeh et al J Res Med Sci. 2010 | 84 (0/84)Premenopausal women with central obesity | This randomized, double-blind, placebo-controlled trial was undertaken in 84 healthy premenopausal women with central obesity. After 2 weeks of run-in on an isocaloric diet, participants were randomly considered to eat hypocaloric diet enriched in legume (HDEL), Arg (5 g/d) and HDEL, selenium (200 µg/d) and HDEL or Arg, selenium and HDEL for 6 weeks. The caloric needs for each subject separately was determined by the equation from the Institute of Medicine, Food, and Nutrition board. All participants in all groups were given a diet of 500 kcal less than their caloric needs in intervention period. | HDEL showed a significant effect in reduction of waist, hip, arm, thigh, calf and breast circumferences, triceps, biceps, subscapular and suprailiac skinfold thicknesses, WHR, SSF, D and EPF. HDEL + Arg + selenium significantly reduced suprailiac skinfold thicknesses; and there was no significant effect of HDEL, Arg, selenium and Arg plus selenium on weight, BMI and fasting NOx. | The study indicates that HDEL + Arg + selenium reduce suprailiac skinfold thicknesses which represents the abdominal obesity reduction. |
| The effects of four hypocaloric diets containing different levels of sucrose or high fructose corn syrup on weight loss and related parameters | Joshua Lowndes Nutr. Jr. 2012 | 162 (35,127)Overweight and Obese Adults | This was a randomized, prospective, double blind trial, with overweight/obese participants measured for body composition and blood chemistry before and after the completion of 12 weeks following a hypocaloric diet All four hypocaloric diets (Groups 1–4) were based on individualized calorie levels using the Mifflin-St Jeor calculation for REE (with activity factor) minus 500 kilocalories (2093 KJ).. | Reductions were observed in all measures of adiposity including body mass, BMI,% body fat, waist circumference and fat mass for all four hypocaloric groups, as well as reductions in the exercise only group for body mass, BMI and waist circumference. | Similar decreases in weight and indices of adiposity are observed when overweight or obese individuals are fed hypocaloric diets containing levels of sucrose or high fructose corn syrup typically consumed by adults in the United States.. |
| Early Effects of a Hypocaloric, Mediterranean Diet on Laboratory Parameters in Obese Individuals | Marta Greco, Mediators of Inflammation,2014 | 23(6/17)Obese pharmacologically treated adults | 23 obese, pharmacologically untreated patients were enrolled and subjected to the determination of anthropometric variables and blood collection at baseline, 1 and 4 months after diet initiation. Mediterranean, hypocaloric (1400–1600 Kcal/d) diet. | Body weight and BMI significantly decreased (P<0.001) after calorie restriction. Weight loss led to an improvement in insulin sensitivity, as indicated by a decrement of both insulin and HOMA-IR index . A more significant variation of insulin in patients achieving a greater body weight loss | A precocious improvement of insulin and leptin sensitivity after a modest calorie restriction and weight reduction |
| Beneficial effects of a High Protein Low Glycemic Load Hypocaloric Diet inoverweight and obese women withpolycystic ovary syndrome:A randomized controlled intervention study | Zohreh Amiri, Journal of the American College of Nutrition,2012 | 60(0/60)Overweight and obese women with PCOS | A total of 60 overweightand obese women with PCOS who did not use insulin-sensitizing agents were recruited and randomlyassigned to 1 of the 2 hypocaloric diet groups for a single-blind clinical trial The groups included a Conventional Hypocaloric Diet (CHCD) (15% of daily energy from protein) and a Modified Hypocaloric Diet(MHCD) with a high-protein, low-glycemic load (30% of daily energy from protein plus low- glycemic-load foods selected from a list) that was prescribed via counseling visits weekly during 12 weeks of study | Weight loss was significant and similar in the 2 groups.Mean of testosterone in the MHCD and CHCD groups decreased from 1.78 ± 0.32 to 1.31 ± 0.26 ng/ml andfrom 1.51 ± 0.12 to 1.15 ± 0.11 ng/ml, respectively (p < 0.001). Follicle sensitizing hormone (FSH),luteinizing hormone (LH), and blood lipids concentrations were not changed except low-density lipoprotein cholesterol (LDL-C) was reduced by 24.5% ± 12.3% p <0.001 for both) after 12 weeks of intervention. MHCD resulted in a significant reduction in insulin level,dhomeostatic model assessment for insulin resistance(HOMA), and high-sensitivity C- reactive protein (hsCRP) concentration (p < 0.001). | Both hypocaloric diets without using any medications and instructed exercise were shown tosignificantly reduce body weight and androgens levels in two groups of women with PCOS. Inaddition a high protein-low GLdiet caused a significant increase in insulin sensitivity and adecrease in hsCRP level when compared to conventional diet. However a study to differentiatethe effects of protein content and glycemic loadin a hypocaloric diet for women with PCOS isneeded |
Conclusion
Obese individuals show a blunted lipid response in response to cholesterol lowering diets. These cholesterol lowering diets do not favourably modify the plasma lipid and lipoproteinin obese individuals. As there is decreased cholesterol excretion and increased cholesterol synthesis which is a result of insulin resistance and inflammation commonly seen in obesity.There is also alteration in the activity of enzymes involved in normal lipid metabolism (eg. LPL) which in turn leads to differential response to change in dietary lipid. On the contrary, a reduction in adipose tissue mass enchances LDL receptor activity by decreasing insulin resistance and inflammation. As a result, weight loss is recommended for overweight/obese individuals to realize the maximal benefits of dietary interventions low in SFA and cholesterol. Hypocaloric diets (500 kcal less from requirement) which bring about weight loss in obese individual have shown to have dual benefit of improving the lipid profile as well. Therefore, in conclusion it can be said that bringing about weight loss first and then improving the lipid profile should be considered while planning dietary interventions.
References
- Shoelson, S.E., Herrero, L., and Naaz, A. Obesity inflammation, and insulin resistance. Gastroenterology, 132:2169–2180 2008.
CrossRef - Schenk S, Saberi M, Olfesky Jm. Insulin sensitivity: modulation by nutrients and inflammation. J Clin Invest,118;2992-3002 2008.
CrossRef - Gylling H, Miettinen TA. Cholesterol absorption, synthesis, and LDLmetabolism in NIDDM. Diabetes Care, 20:90–5 1997.
CrossRef - Berglund L, Lefevre M, Ginsberg HN, Kris-Etherton PM, ElmerPJ,Stewart PW, Ershow A, Pearson TA, Dennis BH, et al. Comparison of monounsaturated fat with carbohydrates as a replacement for saturated fat in subjects with a high metabolic risk profile: studies in thefasting and postprandial states. Am J ClinNutr, 86:1611–20 2007.
- Lichtenstein AH, Matthan NR, Jalbert SM, Resteghini NA, SchaeferEJ, Ausman LM. Novel soybean oils with different fatty acid profilesalter cardiovascular disease risk factors in moderately hyperlipidemicsubjects. Am J ClinNutr, 84:497–504 2006.
- Mustad VA, Etherton TD, Cooper AD, Mastro AM, Pearson TA,Jonnalagadda SS, Kris-Etherton PM. Reducing saturated fat intake isassociated with increased levels of LDL receptors on mononuclearcells in healthy men and women. J Lipid Res, 38:459–68 1997.
- Bronsgeest-Schoute DC, Hermus RJ, Dallinga-Thie GM, Hautvast JG. Dependence of the effects of dietary cholesterol and experimental conditions on serum lipids in man. III. The effect on serum cholesterol of removal of eggs from the diet of free-living habitually egg-eating people.Am J Clin Nutr, 32(11):2193-7 1979 Nov.
- Mukuddem-Petersen J1, Stonehouse Oosthuizen W, Jerling JC, Hanekom SM, White Z. Effects of a high walnut and high cashew nut diet on selected markers of the metabolic syndrome: a controlled feeding trial. Br J Nutr, 97(6):1144-53 2007 June.
CrossRef - Lefevre M1, Champagne CM, Tulley RT, Rood JC, Most MM. Individual variability in cardiovascular disease risk factor responses to low-fat and low-saturated-fat diets in men: body mass index, adiposity, and insulin resistance predict changes in LDL cholesterol. Am J Clin Nutr, 82(5):957-63; quiz 1145-6 2005 Nov.
- Sergio Jansen, Jose Lopez-Miranda,Joaquin Salas, Pedro Castro, Juan A. Paniagua,Fernando Lopez-Segura, Jose M. Ordovas*, Jose A. Jimenez-Pereperez, Angeles Blanco, and Francisco Perez-Jimenez3Plasma Lipid Response to Hypolipidemic Diets in Young Healthy Non-Obese Men Varies with Body Mass Index J. Nutr, vol. 128 no. 7 1144-1149 1998 July 1.
- Hosogai N, Fukuhara A,Oshima K, Miyata Y, Tanaka S, Segawa K,Furukawa S, Tochino Y, Komuro R, et al. Adipose tissue hypoxia in obesity and its impact on adipocytokine dysregulation. Diabetes, 56:901–11 2007.
CrossRef - Stephens JM, Lee J, Pilch PF Tumor necrosis factor-alpha-induced insulin resistance in 3T3–L1 adipocytes is accompanied by a loss of insulin receptor substrate-1 and GLUT4 expression without a loss of insulin receptor-mediated signal transduction. J Biol Chem,272:971–6 1997.
CrossRef - Ruan H, Hacohen N, Golub TR, Van Parijs L, Lodish H . Tumor necrosis factor-alpha suppresses adipocyte-specific genes and activates expression of preadipocyte genes in 3T3–L1 adipocytes: nuclear factor-kappaB activation by TNF-alpha is obligatory. Diabetes,51:1319–36 2002.
CrossRef - Visser M, Bouter LM, McQuillan GM, Wener MH, Harris TB Elevated C-reactive protein levels in overweight and obese adults. JAMA, 282:2131–5 1999.
CrossRef - Sunil K. Kota, Lalit K. Meher Siva K. Kota Sruti Jammul S. V. S. Krishna, Kirtikumar D. Modi, Implications of serum paraoxonase activity in obesity, diabetes mellitus, and dyslipidemiaIndian Journal of Endocrinology and Metabolism / Vol 17 | Issue 3 May-Jun 2013.
- McGillicuddy FC, de la Llera Moya M, Hinkle CC,Joshi MR, Chiquoine EH, Billheimer JT, Rothblat GH, Reilly MP. Inflammation impairs reverse cholesterol transport in vivo. Circulation, 119:1135–45 2009.
CrossRef - Lewis GF, Carpentier A, Adeli K, Giacca . Disordered fat storage and mobilization in the pathogenesis of insulin resistance and type 2 diabetes. Endocr Rev, 23:201–29 2002.
CrossRef - Mekki N, Christofilis MA, Charbonnier M, Atlan-Gepner C, Defoort C, Juhel C, Borel P, Portugal H, Pauli AM, et al. Influence of obesity and body fat distribution on postprandial lipemia and triglyceride-rich lipoproteins in adult women. J Clin Endocrinol Metab, 84:184–91 1999.
CrossRef - Tobin KA, Ulven SM, Schuster GU, Steineger HH, Andresen SM, Gustafsson JA, Nebb HI Liver X receptors as insulin-mediating factors in fatty acid and cholesterol biosynthesis. J Biol Chem, 277:10691–7 2002.
CrossRef - Lefevre M, Champagne CM, Tulley RT, Rood JC, Most MM. Individual variability in cardiovascular disease risk factor responses to low-fat and low-saturated-fat diets in men: body mass index, adiposity, and insulin resistance predict changes in LDL cholesterol. Am J Clin Nutr, 82:957–63 2005.
- Yu L, York J, von Bergmann K, Lutjohann D, Cohen JC, Hobbs H. Stimulation of cholesterol excretion by the liver X receptor agonist requires ATP-binding cassette transporters G5 and G8. J Biol Chem,278:15565–70 2003.
CrossRef - Tsubakio-Yamamoto K,Nishida M, Nakagawa-Toyama Y, Masuda D, Ohama T, Yamashita S. Current therapy for patients with sitosterolemia–effect of ezetimibe on plant sterol metabolism. J Atheroscler Thromb, 17:891–900 2010.
CrossRef - Klett EL, Lee MH, Adams DB, Chavin KD,Patel SB. Localization of ABCG5 and ABCG8 proteins in human liver, gall bladder and intestine. BMC Gastroenterol, 4:21 2004.
CrossRef - Berge KE, Tian H, Graf GA, Yu L, Grishin NV, Schultz J, Kwiterovich P, Shan B, Barnes R, et al Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters.Science,290:1771–5 2000.
CrossRef - Santosa S, Demonty I, Lichtenstein AH, Ordovas JM, Jones PJ. Single nucleotide polymorphisms in ABCG5 and ABCG8 are associated with changes in cholesterol metabolism during weight loss. J Lipid Res, 48:2607–13 2007.
CrossRef - Li T, Ma H, Chiang JY. TGFβ1, TNFα, and insulin signaling crosstalk in regulation of the rat cholesterol 7α-hydroxylase gene expression. J Lipid Res,49:1981–9 2008.
CrossRef - Katcher HI1, Legro RS, Kunselman AR, Gillies PJ, Demers LM, Bagshaw DM, Kris-Etherton PM. The effects of a whole grain-enriched hypocaloric diet on cardiovascular disease risk factors in men and women with metabolic syndrome, Am J Clin Nutr, 87(1):79-90 2008 Jan.
- Alizadeh M1, Daneghian S, Ghaffari A, Ostadrahimi A, Safaeiyan A, Estakhri R, Gargari BP.The effect of hypocaloric diet enriched in legumes with or without L-arginine and selenium on anthropometric measures in central obese women. Am J Clin Nutr, 87(1):79-90 2008 Jan.
- Joshua Lowndes1, Diana Kawiecki1, Sabrina Pardo1, Von Nguyen1, Kathleen J Melanson2, Zhiping Yu1 and James M Rippe1* The effects of four hypocaloric diets containing different levels of sucrose or high fructose corn syrup on weight loss and related parameters,Nutrition Journal, 11:55 doi:10.1186/1475-2891-11-55 2012.
CrossRef - Marta Greco,1 Eusebio Chiefari,1 Tiziana Montalcini,2 Francesca Accattato,1 Francesco S. Costanzo,3 Arturo Pujia,2 Daniela Foti,1 Antonio Brunetti,1 and Elio Gulletta1, Early Effects of a Hypocaloric, Mediterranean Diet on Laboratory Parameters in Obese Individuals, Mediators of Inflammation,Volume 2014 , Article ID 750860, 8 pages 2014.
- Mehrabani HH1, Salehpour S, Amiri Z, Farahani SJ, Meyer BJ, Tahbaz F. Beneficial effects of a high-protein, low-glycemic-load hypocaloric diet in overweight and obese women with polycystic ovary syndrome: a randomized controlled intervention study,J Am Coll Nutr, 31(2):117-25 2012 Apr.
CrossRef.

This work is licensed under a Creative Commons Attribution 4.0 International License.








