

Introduction
Soy-drink attracts consumers as a substitute for dairy-based food products because of its protein content and lack of cholesterol and lactose. It is also an excellent source of high protein, vitamin B2, vitamin B12, potassium, and isoflavones. Soy-drink can be regarded as an alternative for bovine milk, including an ideal nutritional portion for lactose-intolerant populations, and has key components for the successful growth of lactic acid bacteria (LAB)1. Prebiotics are specific food types that are non-digestible but are considered a significant element as a growth factor for lactic acid bacteria2. One of the most frequently employed prebiotic is inulin, a fructan of non-digestible nature that possesses a specific mix of broad nutritional aspects, including its technical advantages. Inulin, when incorporated into yogurt, shows its efficacy as a bulking agent, the substitution of fats, lower-calorie sweeteners, and modification of textures, thereby predominantly changing the organoleptic perception fermented food products3. It is believed that soy drink supplementation with skimmed milk powder (SMP) promotes nutritional parameters and associated health applications, including the development of active soy aglycones from glucosidic conjugates. SMP also enhances the rate of growth of starter culture in the yogurt, which could, in turn, act as a source of active isoflavone aglycones infused with richer protein as well as calcium content4. Whey protein concentrate (WPC 70, 70 gm of protein per 100gm) is generally considered as a source of protein that has the efficacy for uniform mixing up with different fermented and non-fermented dairy foods due to their rich nutritional impact and bio-functional properties5. WPCs are capable of induction of textural features of dairy foods, and they possess sufficient concentrations of lactose that serve as a matrix where biosynthesis of fermentable sugars occurs by the action of lactic acid bacteria5.
Amongst lactic acid bacteria, β-D-Glucosidase activity is more specifically popular in Lactobacillus spp. since it contributes to the dietary/organoleptic attributes of fermented dairy foods. Through deglucosylation, flavor elements are released from the precursors that had undergone glucosylation and are capable of raising the bioavailability of health benefitting metabolites along with toxic dietary compounds6. Soy-drink consumption by masses suffers major hindrance due to their typical beany flavor7. This flavor is due to the presence of various oligosaccharides (non-digestible), causing flatulence, bloating, abdominal pain, and other gastrointestinal disorders. These problems could be mitigated if lactic acid bacteria supplemented with different nutritional factors in soy-drink7. In correlation with the above-stated enzyme activities, the proteolytic activity of Lactobacillus spp. increases the physiological activity of cultured dairy products by liberating several biologically active peptides8. Milk has always been proved to be a source of free amino acids and peptides, making it a rich medium for LAB to grow efficiently9. This group of bacteria constitutes proteinase as well as peptidase, which could synthesize casein derived amino acids that play an essential role in organic nitrogen source10. The study was commenced for determining and comparing the effect of few nutritional supplements (inulin, WPC 70, SMP) on the growth behavior, β-glucosidase, β-galactosidase, and proteolytic activities of indigenous Lactobacillus in soy-drink.
Materials and Methods
Bacterial Strains
The Lactobacillus cultures bearing NCBI GenBank accession numbers, namely L. fermentum K3AA (KU644575), L. rhamnosus K4EE (KX950834), L. fermentum K5 (KU213668) L. fermentum K7 (KU213665), L. helveticus K14 (KU644578), L. fermentum K16 (KU213667), L. acidipiscis 27A (KY234394), L. plantarum RD7 (MF155569.1) were obtained from the culture collection of Animal Science Laboratory, Department of Rural Development and Agricultural Production, North-Eastern Hill University, Tura Campus, Meghalaya, India11. These selected strains were employed due to their rich probiotic potentiality, studied previously12.
Soybean Sample
The local variety of soybeans (Rymbai-ktung in local language) were acquired from a local marketplace in Tura town of West Garo Hills district in Meghalaya. Meghalaya’s tribal population mainly uses this variety of soybean to prepare their various traditional fermented soy foods13.
Preparation of Inoculum
The purely MRS (De Man, Rogosa and Sharpe) grown Lactobacillus isolates were first transferred into sterile rehydrated skim milk followed by preserving them in glycerol stocks by placing them in 1 mL aliquots in cryovials and further storing those at −40°C. Each of the Lactobacillus cultures was inoculated into a skimmed milk medium (HiMedia, Mumbai, India) and incubated at 37℃ for another 18 h after two consecutive transfers of the lactic strains in MRS broth, which were incubated at 37℃ for 24 h. Finally, the lactic strains were inoculated into soy-drink14.
Soy-Drink Preparation
The preparation of soy-drink was achieved by the method adapted from Hati et al.14. Around 2.5 L of soy-drink was obtained per 500 g of soybeans and was autoclaved at 121℃ for 20 min. followed by cooling to 42℃. The soy-drink was divided into equal parts with different nutritional treatments viz. inulin, WPC 70 and SMP at the rate of 2%. All of the samples were stored in refrigeration conditions until further analysis (6℃-8℃).
Evaluation of pH, Acidity, and Lactobacillus Counts in Soy Drink Supplemented with Nutritional Factors
All the Lactobacillus cultures were activated by growing in sterilized soy-drink. Each of the cultures was added in soy-drink to equivalent levels with different nutritional treatments viz. inulin, WPC 70, and SMP at the rate of 2%. After mixing them thoroughly, the samples were incubated at 37℃ for different time intervals of 0, 3, 6, 9, 12, and 24h. After each time interval, samples were obtained to evaluate pH, titratable acidity, and viable lactic cell counts. pH of the samples was estimated by using a digital pH meter (OAKTON pH700, India). The titratable acidity was measured by the procedure described by Indian Standards15,21. Lactobacilli counts for each of the isolates from different treatments were checked in deMan Rogosa and Sharpe (MRS) Agar was determined as per the method described by IDF standards16,21. The colonies were calculated, and the counts were expressed as log CFU/ml.
Proteolytic Activity of Lactobacillus Cultures in Soy Drink Supplemented with Nutritional Factors
Soy-drink was supplemented with inulin, SMP and WPC 70 at 2% level each and the proteolytic activity of the Lactobacillus cultures engaged in each supplementation were analyzed. An aliquot of 2.5 ml from each lactobacilli strains in soy drink supplemented by various nutritional factors was mixed with 0.75% trichloroacetic acid (TCA) allowed to stand for 10 minutes. Using the Whatman filter paper 42, the mix was filtered. In a clean test tube, 200 μl of the obtained filtrate was applied to 3 ml of o-phthaldialdehyde (OPA) reagent and mixed uniformly, followed by incubation for 2 minutes at room temperature. The peptides emitted in soy milk under different nutritional treatments were measured as free amino acid absorption at 340 nm by using Double beam Spectrophotometer 2202S, Systronics Ltd., India, following the OPA method of Donkor et al.17. Proteolytic activity was evaluated at 340 nm as absorbance of free amino acid groups, after 24 h of fermentation at 37℃. All the analysis was carried out in triplicates.
β-Glucosidase Activity of Lactobacillus Cultures in Soy Drink Supplemented with Nutritional Factors
The β-glucosidase activity of the Lactobacillus cultures incorporated in soy-drink supplemented with inulin, SMP and WPC 70 at 2% level were interpreted by analyzing the hydrolysis rate of p-nitrophenol β-D-glucopyranoside (pNPG) (HiMedia, India). The method was adapted from Otieno et al.18 18 and Hati et al.19. 5mM of p-nitrophenyl -D-glucopyranoside (substrate) was prepared in a 100mM sodium phosphate buffer with a pH of 7.0. Subsequently, 500 μl of this substrate was added to 5 ml of each aliquot sample (of different nutritional supplementation) and incubated at 37°C for 30 min. 250 μl of chilled 0.2 M sodium carbonate was implemented to terminate the reaction. At 15000 rpm for 30 minutes, the mix acquired was centrifuged and filtered through a syringe filter (0.45μ). Using a UV-Vis double beam spectrophotometer at 410 nm, the amount of p-nitrophenol released was evaluated spectrophotometrically.
β-Galactosidase Activity of Lactobacillus Cultures in Soy Drink Supplemented with Nutritional Factors
The β-galactosidase enzyme assay was determined for the Lactobacillus cultures incorporated in soy-drink supplemented with inulin, SMP and WPC 70 at 2% level employing cell disruption methodologies adapted from Prasad et al.20 and Makwana et al.21 with few modifications.
Extraction of Enzyme
The Lactobacillus cultures were added at 2% rate in soy milk supplemented with inulin, SMP and WPC 70, and the culture flasks were incubated at 37°C for 24 hours, followed by centrifugation of cells at 5000 rpm for 15 minutes at 4°C for supernatant extraction, which is considered to comprise extracellular enzymes. The cell pellet was crushed by intense vortexing and washed twice with a 0.05 M sodium phosphate buffer (pH 6.8) and centrifuged for 15 min at 4°C at 5000 rpm, followed by resuspending the washed pellets in 10 mL of 0.05 M phosphate buffer for intracellular enzyme extraction using the methods of cell disruption as prescribed below.
Sonication
The cell suspensions were subjected to sonication by fixing up a few parameters viz. Pulse-30 seconds on /30 seconds off and 60% amplitude in an ice bath. The extract was further subjected to centrifugation at 5000 rpm at 4°C for 15 min after sonication. The supernatant with the crude enzyme extract was stored at -20°C before further enzymatic assays were used.
Enzyme Assay
The β-galactosidase activity was evaluated by supplementing nutritional elements with each Lactobacillus isolate in soy milk. The reaction mixture consisted of 0.5 mL of extracted enzyme-containing supernatant and 2.0 mL of 15 mM O-nitrophenyl β-D-galactopyranoside (ONPG) in a sodium phosphate buffer of 0.05 M (pH 6.8). 0.5 mL of 0.1 M sodium carbonate was applied to the mixture after incubation for 20 min at 37°C to halt the reaction. With a UV-Vis double beam spectrophotometer, the absorbance was read at 420 nm.
Statistical Analysis
Data presented in the study are the average of three independent replicates, and the results are expressed as mean ± standard deviation. Two-way analysis of variance (ANOVA) was applied, and the post-hoc comparison was acquired by Tukey’s HSD test with the least significant difference of p≤0.05 using IBM SPSS Statistical program (Ver. 20).
Results and Discussions
Growth Behavior of Lactobacillus Cultures in Soy-Drink with Nutritional Factors
The tendency to produce flatulence and other related disorders in the gastrointestinal tract due to soy-drink consumption can be resolved if the various oligosaccharides (specifically stachyose and raffinose) are utilized Lactobacillus cultures 14. This will be accomplished by fermentation with additional nutritional supplements thereby improving the overall digestion. pH, titratable acidity and viable lactic bacteria counts were determined after various time periods viz. 0, 3, 6, 9, 12 and 24 h of soy-drink fermentation at 37°C supplemented with inulin (Table 1), WPC 70 (Table 2) and SMP (Table 3), at the rate of 2%.
The maximum lowering of pH was observed in L. acidipiscis K27A at 2% inulin (ranging from 6.15 to 3.92) followed by L. fermentum K16 at 2% WPC 70 (ranging from 6.32 to 3.52). Similarly, a gradual drop in pH was noticed in L. helveticus K14 at 2% SMP supplementation to soy-drink ranging from 6.38 to 3.59 after 24h incubation. The pH reduction was rapid in all the eight Lactobacillus cultures, especially for L. acidipiscis K27A and L. helveticus K14 throughout the incubation period. This may be because of soy protein’s low buffering ability compared to related milk proteins in soy-drink22. Therefore, supplementation of the nutritional factors appears to have played a primary role in reducing pH significantly (p≤0.05) in soy-drink by adding more nutrition to the lactic bacterial cultures. Although secondary products, as such the butter flavor substances like diacetyl are also produced but as an index for titratable acidity, the lactic acid content is primarily held as a significant parameter for fermentation of yogurt23.
In this study, the highest production of lactic acid (%) was seen in L. fermentum K5 at 2% inulin supplementation, ranging from 0.132 to 0.481. Similarly, L. fermentum K16 showed the highest % of lactic acid production at 2% WPC 70 supplementation ranging from 0.128 to 0.468. L. helveticus K14 also showed the highest % of lactic acid production at 2% SMP supplementation, significantly ranging from 0.131 to 0.441 after 24h incubation. Few of the LAB has a low growth rate in the soy-drink due to the absence of lactose and hence supplementation by combining various nutritional supplements and probiotics could bring out the staggering health benefits24. From previous studies, it was stated in a study strain of Lactobacillus delbrueckii spp. bulgaricus was recommended to be employed for fermenting soy-drink with the addition of inulin since the strain had the potential to generate higher concentrations of lactic acid than other lactobacilli used in the study25. Kothari26 also reported that even after enriching with lactose, the lactic acid production from soy-drink was held insufficient for manufacturing fermented products, but the crisis was resolved with whey protein concentrate. Similarly, after the addition of skim milk (50:50). Rajasekaran and Rajor27 observed a decrease of soy solids, thereby producing a desirable yogurt-like product.
The highest lactic bacterial counts were exhibited by L. plantarum RD7 at 2% inulin (8.58 ± 0.033 log CFU/mL) supplementation followed by L. acidipiscis K27A at 2% WPC 70 (8.50 ± 0.022 log CFU/mL). L. plantarum RD7 also showed high cell count at 2% SMP (8.47 ± 0.038 log CFU/mL) supplementation to soy-drink after a period of 24h. The study stands in complete agreement with Hati et al.28 where the strains L. bulgaricus NCDC and L. helveticus MTCC 5463 cultures proved to be predominant compared to the other cultures by exhibiting high lactic bacterial cell count at 2% WPC 70 supplemented in soy-drink. Similarly, lactic acid production and growth rate were lower as a comparison to fermented milk since soy-drink is lactose deficient, and hence lactic acid bacteria were unable to produce acidity compared to yogurt made from cow milk29. Supplementing with WPC provided with added nutrition to the Lactobacillus cultures throughout the growth period. Moreover, in order to enhance the growth of lactic acid bacteria, inulin and whey protein concentrate are considered essential elements for soy-drink to be supplemented30.
Table 1: Impact of Inulin in pH, Acidity and Lactic Bacterial Counts during Soy-Drink Fermentation
| Conc. of Inulin | Strains | |||||||
| Fermentation period (in hours) | ||||||||
| 0 | 3 | 6 | 9 | 12 | 24 | |||
| pH | 2% | K3A | 6.44 ± 0.12 a | 6.18 ± 0.05 a | 5.85 ± 0.12 b | 5.51 ± 0.17 b | 5.22 ± 0.12 b | 4.49 ± 0.14 c |
| K4E | 6.36 ± 0.11 a | 6.20 ± 0.16 a | 6.08 ± 0.13 a | 5.76 ± 0.09 b | 5.55 ± 0.10 b | 4.27 ± 0.20 c | ||
| K5 | 6.29 ± 0.14 a | 6.08 ± 0.11 a | 5.75 ± 0.09 b | 5.48 ± 0.10 b | 5.07 ± 0.14 b | 4.15 ± 0.02 c | ||
| K7 | 6.25 ± 0.08 a | 6.08 ± 0.16 a | 5.65 ± 0.05 b | 5.11 ± 0.17 b | 4.71 ± 0.22 c | 4.05 ± 0.15 c | ||
| K14 | 6.14 ± 0.12 a | 6.02 ± 0.17 a | 5.63 ± 0.10 b | 5.73 ± 0.08 b | 5.24 ± 0.17 b | 4.57 ± 0.03 c | ||
| K16 | 6.22 ± 0.19 a | 6.12 ± 0.13 a | 5.58 ± 0.24 b | 5.27 ± 0.17 b | 4.71 ± 0.11 c | 4.00 ± 0.18 c | ||
| K27A | 6.15 ± 0.11 a | 5.75 ± 0.14 b | 5.59 ± 0.19 b | 5.17 ± 0.21 b | 4.62 ± 0.19 c | 3.92 ± 0.02 d | ||
| RD7 | 6.28 ± 0.09 a | 6.13 ± 0.11 a | 5.55 ± 0.08 b | 5.37 ± 0.18 b | 4.80 ± 0.07 c | 4.11 ± 0.08 d | ||
| TA | 2% | K3A | 0.118 ±0.14b | 0.133 ±0.04b | 0.175 ±0.10b | 0.250 ±0.18 c | 0.331 ±0.12 d | 0.405 ±0.16e |
| K4E | 0.124 ±0.03b | 0.192 ±0.20b | 0.258 ±0.01 c | 0.322 ±0.14d | 0.368 ±0.08 d | 0.413 ±0.11e | ||
| K5 | 0.132 ±0.11b | 0.156 ±0.18b | 0.222 ±0.12c | 0.264 ±0.02 c | 0.390 ±0.23 d | 0.481 ±0.22e | ||
| K7 | 0.125 ±0.12b | 0.145 ±0.20b | 0.168 ±0.18b | 0.212 ±0.06 c | 0.268 ±0.19 c | 0.368 ±0.20d | ||
| K14 | 0.120 ±0.06b | 0.141 ±0.14b | 0.165 ±0.09b | 0.209 ±0.11 c | 0.255 ±0.15 c | 0.314 ±0.25d | ||
| K16 | 0.128 ±0.11b | 0.156 ±0.16b | 0.191 ±0.18b | 0.249 ±0.18 c | 0.308 ±0.11 d | 0.365 ±0.17d | ||
| K27A | 0.131 ±0.08b | 0.161 ±0.01b | 0.186 ±0.20b | 0.269 ±0.13 c | 0.339 ±0.22 d | 0.401 ±0.14e | ||
| RD7 | 0.120 ±0.15b | 0.145 ±0.22b | 0.184 ±0.04b | 0.227 ±0.24 c | 0.343 ±0.16 d | 0.471 ±0.17e | ||
| VLC | 2% | K3A | 6.75 ±0.018a | 6.90 ±0.026a | 7.20 ±0.028b | 7.36 ±0.066b | 7.66 ±0.035 b | 8.02 ±0.093c |
| K4E | 7.11 ±0.033b | 7.26 ±0.044b | 7.48 ±0.078b | 7.70 ±0.101b | 7.96 ±0.047 b | 8.38 ±0.025c | ||
| K5 | 6.40 ±0.066a | 6.77 ±0.014a | 6.90 ±0.022a | 7.21 ±0.016b | 7.48 ±0.077 b | 8.09 ±0.022c | ||
| K7 | 6.82 ±0.021a | 7.06 ±0.052b | 7.25 ±0.050b | 7.42 ±0.052b | 7.68 ±0.093 b | 8.11 ±0.098c | ||
| K14 | 6.77 ±0.026a | 6.92 ±0.033a | 7.21 ±0.100b | 7.41 ±0.065b | 7.85 ±0.100 b | 8.35 ±0.015c | ||
| K16 | 6.32 ±0.077a | 6.70 ±0.044a | 6.93 ±0.020a | 7.12 ±0.038b | 7.44 ±0.017 b | 7.97 ±0.201b | ||
| K27A | 6.55 ±0.025a | 6.81 ±0.066a | 7.03 ±0.017b | 7.22 ±0.030b | 7.39 ±0.028 b | 8.08 ±0.047c | ||
| RD7 | 7.01 ±0.021b | 7.25 ±0.110b | 7.41 ±0.066b | 7.62 ±0.025b | 8.05 ±0.025 c | 8.58 ±0.033c | ||
TA-Titratable Acidity (%), VLC-Viable lactic bacterial count (log10CFU/mL). Values are represented in mean ± SD with three independent determinations (n = 3) from each sample. Values bearing different superscripts in each cell differ significantly from each other, as evidenced by Tukey’s test at p ≤ 0.05.
Table 2: Impact of WPC 70 in pH, Acidity and Lactic Bacterial Counts during Soy-Drink Fermentation
| Conc. of Inulin | Strains | |||||||
| Fermentation period (in hours) | ||||||||
| pH | 2% | 0 | 3 | 6 | 9 | 12 | 24 | |
| K3A | 6.25 ± 0.02 a | 5.68 ± 0.03 b | 5.35 ± 0.09 b | 5.08 ± 0.07 b | 4.51 ± 0.02 c | 4.09 ± 0.02 c | ||
| K4E | 6.32 ± 0.03 a | 6.02 ± 0.06 a | 5.68 ± 0.10 b | 5.17 ± 0.05 b | 4.55 ± 0.05 c | 3.97 ± 0.03 d | ||
| K5 | 6.19 ± 0.04 a | 5.78 ± 0.03 b | 5.25 ± 0.09 b | 4.77 ± 0.08 c | 4.11 ± 0.04 c | 3.85 ± 0.04 d | ||
| K7 | 6.29 ± 0.07 a | 5.78 ± 0.06 b | 5.25 ± 0.12 b | 4.61 ± 0.07 c | 4.21 ± 0.03 c | 3.95 ± 0.05 d | ||
| K14 | 6.24 ± 0.08 a | 5.66 ± 0.01 b | 5.33 ± 0.10 b | 4.73 ± 0.04 c | 4.34 ± 0.07 c | 3.88 ± 0.03 d | ||
| K16 | 6.32 ± 0.09 | 5.71 ± 0.02 b | 5.28 ± 0.02 b | 4.52 ± 0.02 c | 4.01 ± 0.03 c | 3.52 ± 0.01 d | ||
| K27A | 6.45 ± 0.10 a | 6.05 ± 0.12 a | 5.69 ± 0.09 b | 4.97 ± 0.01 c | 4.42 ± 0.01 c | 3.89 ± 0.02 d | ||
| RD7 | 6.66 ± 0.05 a | 6.28 ± 0.11 a | 5.75 ± 0.04 b | 5.07 ± 0.02 b | 4.65 ± 0.07 c | 4.09 ± 0.04 c | ||
| TA | 2% | K3A | 0.166 ±0.04a | 0.170 ±0.04 a | 0.195 ±0.07 a | 0.221 ±0.08 b | 0.301 ±0.04 c | 0.415 ±0.06 d |
| K4E | 0.144 ±0.03a | 0.152 ±0.20 a | 0.178 ±0.01 a | 0.235 ±0.04 b | 0.318 ±0.03 c | 0.423 ±0.07 d | ||
| K5 | 0.127 ±0.01a | 0.136 ±0.08 a | 0.147 ±0.02 a | 0.214 ±0.02 b | 0.298 ±0.01 b | 0.445 ±0.12 d | ||
| K7 | 0.135 ±0.02a | 0.141 ±0.15 a | 0.160 ±0.12 a | 0.232 ±0.01 b | 0.305 ±0.09 c | 0.423 ±0.10 d | ||
| K14 | 0.118 ±0.04a | 0.126 ±0.07 a | 0.135 ±0.06 a | 0.198 ±0.09 a | 0.287 ±0.05 b | 0.404 ±0.01 d | ||
| K16 | 0.128 ±0.08a | 0.138 ±0.11 a | 0.151 ±0.08 a | 0.199 ±0.07 a | 0.299 ±0.07 b | 0.468 ±0.06 d | ||
| K27A | 0.110 ±0.09a | 0.121 ±0.01 a | 0.136 ±0.10 a | 0.178 ±0.03 a | 0.278 ±0.10 b | 0.447 ±0.09 d | ||
| RD7 | 0.119 ±0.10a | 0.130 ±0.02 a | 0.144 ±0.04 a | 0.187 ±0.04 a | 0.265 ±0.06 b | 0.411 ±0.07 d | ||
| VLC | 2% | K3A | 6.55 ±0.014a | 6.76 ±0.052 a | 6.95 ±0.028 a | 7.31 ±0.067 b | 7.62 ±0.031 b | 8.14 ±0.088 c |
| K4E | 7.05 ±0.037b | 7.18 ±0.079 b | 7.45 ±0.093 b | 7.62 ±0.032 b | 7.77 ±0.058 b | 8.23 ±0.025 c | ||
| K5 | 6.52 ±0.064a | 6.77 ±0.014 a | 7.90 ±0.027 b | 8.15 ±0.093 c | 8.28 ±0.079 c | 8.78 ±0.066 c | ||
| K7 | 6.71 ±0.021a | 6.92 ±0.055 a | 7.35 ±0.025 b | 7.91 ±0.052 b | 7.48 ±0.038 b | 7.91 ±0.093 b | ||
| K14 | 6.83 ±0.093a | 7.12 ±0.087 b | 7.41 ±0.032 b | 7.79 ±0.066 b | 7.80 ±0.067 b | 8.17 ±0.014 c | ||
| K16 | 6.77 ±0.074a | 6.90 ±0.047 a | 7.22 ±0.060 b | 7.45 ±0.031 b | 7.24 ±0.055 b | 7.77 ±0.052 b | ||
| K27A | 7.11 ±0.024b | 7.25 ±0.064 b | 7.43 ±0.021 b | 7.72 ±0.028 b | 8.19 ±0.028 c | 8.50 ±0.022 c | ||
| RD7 | 6.71 ±0.028a | 6.95 ±0.088 a | 7.18 ±0.064 b | 7.32 ±0.022 b | 7.45 ±0.022 b | 8.01 ±0.034 c | ||
TA-Titratable Acidity (%), VLC-Viable lactic bacterial count (log10CFU/mL). Values are represented in mean ± SD with three independent determinations (n = 3) from each sample. Values bearing different superscripts in each cell differ significantly from each other, as evidenced by Tukey’s test at p ≤ 0.05.
Table 3: Impact of SMP in pH, Acidity and Lactic Bacterial Counts during Soy-Drink Fermentation
| Conc. of Inulin | Strains | |||||||
| Fermentation period (in hours) | ||||||||
| 0 | 3 | 6 | 9 | 12 | 24 | |||
| pH | 2% | K3A | 6.25 ±0.04a | 6.00 ± 0.08 a | 5.75 ± 0.09 b | 5.46 ± 0.02 b | 5.22 ± 0.10 b | 4.32 ± 0.10 c |
| K4E | 6.18 ± 0.11 a | 5.88 ± 0.10 b | 5.59 ± 0.11 b | 5.32 ± 0.07 b | 5.05 ± 0.04 b | 4.44 ± 0.08 c | ||
| K5 | 6.15 ± 0.05 a | 5.92 ± 0.07 b | 5.62 ± 0.07 b | 5.39 ± 0.07 b | 5.17 ± 0.11 b | 4.33 ± 0.01 c | ||
| K7 | 6.10 ± 0.01 a | 5.85 ± 0.05 b | 5.43 ± 0.09 b | 5.04 ± 0.12 b | 4.75 ± 0.04 c | 3.90 ± 0.15 d | ||
| K27A | 6.20 ± 0.04 a | 6.04 ± 0.07 a | 5.78 ± 0.05 b | 5.40 ± 0.07 b | 5.11 ± 0.05 b | 4.11 ± 0.07 c | ||
| K16 | 6.30 ± 0.10 a | 6.12 ± 0.04 a | 5.74 ± 0.09 b | 5.48 ± 0.05 b | 4.88 ± 0.04 c | 3.72 ± 0.04 d | ||
| K14 | 6.38 ± 0.10 a | 5.89 ± 0.10 b | 5.41 ± 0.04 b | 5.10 ± 0.12 b | 4.65 ± 0.09 c | 3.59 ± 0.02 d | ||
| RD7 | 6.26 ± 0.09 a | 6.01 ± 0.11 b | 5.75 ± 0.07 b | 5.38 ± 0.02 b | 4.85 ± 0.02 c | 3.77 ± 0.04 d | ||
| TA | 2% | K3A | 0.120 ±0.02 a | 0.145 ±0.06 a | 0.161 ±0.01 a | 0.196 ±0.02 b | 0.228 ±0.09 b | 0.321 ±0.05 c |
| K4E | 0.135 ±0.05 a | 0.158 ±0.10 a | 0.188 ±0.08 a | 0.233 ±0.05 b | 0.251 ±0.07 b | 0.402 ±0.06 d | ||
| K5 | 0.119 ±0.03 a | 0.136 ±0.03 a | 0.162 ±0.11 a | 0.200 ±0.13 b | 0.233 ±0.04 b | 0.433 ±0.05 d | ||
| K7 | 0.122 ±0.10 a | 0.148 ±0.07 a | 0.175 ±0.09 a | 0.206 ±0.08 b | 0.237 ±0.08 b | 0.420 ±0.08 d | ||
| K14 | 0.131 ±0.07 a | 0.162 ±0.05 a | 0.191 ±0.02 a | 0.229 ±0.13 b | 0.255 ±0.09 b | 0.441 ±0.07 d | ||
| K16 | 0.119 ±0.05 a | 0.136 ±0.02 a | 0.151 ±0.11 a | 0.189 ±0.10 a | 0.228 ±0.06 b | 0.399 ±0.03 d | ||
| K27A | 0.128 ±0.09 a | 0.143 ±0.07 a | 0.169 ±0.10 a | 0.198 ±0.03 b | 0.239 ±0.11 b | 0.409 ±0.06 d | ||
| RD7 | 0.138 ±0.08 a | 0.165 ±0.06 a | 0.197 ±0.06 b | 0.242 ±0.07 b | 0.266 ±0.09 b | 0.431 ±0.01 d | ||
| VLC | 2% | K3A | 6.41 ±0.020 a | 6.78 ±0.025 a | 7.12 ±0.030 b | 7.34 ±0.043 b | 7.63 ±0.044 b | 8.06 ±0.091 c |
| K4E | 7.00 ±0.036 b | 7.26 ±0.091 b | 7.41 ±0.110 b | 7.62 ±0.100 b | 7.93 ±0.042 b | 8.39 ±0.020 c | ||
| K5 | 6.75 ±0.028 a | 6.95 ±0.015 a | 7.30 ±0.042 b | 7.51 ±0.036 b | 7.70 ±0.035 b | 8.29 ±0.100 c | ||
| K7 | 6.68 ±0.016 a | 7.14 ±0.050 b | 7.33 ±0.042 b | 7.61 ±0.050 b | 7.88 ±0.091 b | 8.31 ±0.025 c | ||
| K14 | 7.10 ±0.033 b | 7.28 ±0.025 b | 7.51 ±0.088 b | 7.72 ±0.011 b | 8.01 ±0.018 b | 8.36 ±0.016 c | ||
| K27A | 6.55 ±0.070 a | 6.81 ±0.038 a | 7.13 ±0.033 b | 7.30 ±0.016 b | 7.56 ±0.010 b | 8.10 ±0.105 c | ||
| K16 | 6.42 ±0.055 a | 6.61 ±0.068 a | 7.05 ±0.100 b | 7.25 ±0.033 b | 7.46 ±0.017 b | 8.18 ±0.044 c | ||
| RD7 | 7.09 ±0.022 b | 7.24 ±0.100 b | 7.48 ±0.025 b | 7.70 ±0.025 b | 8.10 ±0.025 c | 8.47 ±0.038 c | ||
TA-Titratable Acidity (%), VLC-Viable lactic bacterial count (log10CFU/mL). Values are represented in mean ± SD with three independent determinations (n = 3) from each sample. Values bearing different superscripts in each cell differ significantly from each other, as evidenced by Tukey’s test at p ≤ 0.05.
Proteolytic Activity of Lactobacillus Cultures in Soy-Drink with Nutritional Factors
Amino acids and production of peptides resulting from the biosynthesis of milk proteins by the enzymatic activity of LAB and complete synthesis of the essential amino acids are considered as a crucial metabolic mechanism of LAB31. The extent of proteolytic activity at the time during fermentation of soy-drink was measured by determining the free amino groups employing the o-phthaldialdehyde (OPA) method17. The soy-drink was supplemented with inulin, SMP and WPC 70 at 2% level and the proteolytic activity of the eight Lactobacillus cultures was analyzed, and plain soy-drink without any nutritional supplementation was considered as control. Proteolytic activity was evaluated after a time interval of 24 h of fermentation at 37°C. L. helveticus K14 showed maximum proteolysis at 2% levels of inulin and SMP addition in soy-drink compared to the rest of the cultures. L. fermentum K7 also showed the maximum proteolytic activity at 2% level of WPC 70 addition in soy-drink as depicted in Figure 1.
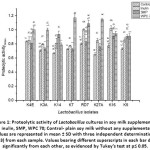 |
Figure 1: Proteolytic activity of Lactobacillus cultures in soy milk supplemented with inulin, SMP, WPC 70; Control- plain soy milk without any supplementation. |
These differences in the amount of amino groups liberated during soy-drink fermentation with additional nutritional supplementation could probably be related to the different proteinases and peptidases of the eight Lactobacillus strains, which appeared to be strain-dependent9 with different nutritional treatments. Hence, from the results of univariate analysis of variance, it was derived that proteolytic activity in all the eight cultures was significantly highest (p≤0.05) after supplementation with WPC 70 as compared to inulin and SMP. The extent of proteolysis with different soy-drink treatments was significantly different (p≤0.05) for all batches infused with inulin, WPC 70 and SMP. From the above-stated results, it was witnessed that the range of proteolysis differed amongst the Lactobacillus cultures that were studied and proved to be time and strain dependant17. With relation to our study, maximum proteolysis was shown by Streptococcus thermophilus MTCC 5462, L. rhamnosus MTCC 5945, and L. helveticus MTCC 5463 at 1.5% and 2% levels of WPC addition in soy-drink in comparison to the rest of lactic bacterial cultures after 24 h of incubation28. Hati et al.32 also testified that L. rhamnosus C6 was predominant with high proteolytic activity as 565.83 lg serine/mL in soy-drink. Probiotic strain L. helveticus M92 exhibited maximum proteolysis with WPC enriched soy-drink33. L. rhamnosus C6 was claimed as rich proteolytic bacteria34 after fermenting the soy-drink from the qualitative/quantitative results in their study. From the above study, it can thereby be derived that whey protein concentrate has the potential of successfully substituting skimmed milk powder for supplementing total solids and fermentable sugars in soy-drink for yogurt development and manufacture5.
β-Glucosidase Activity of Lactobacillus Cultures in Soy-Drink with Nutritional Factors
A total of eight indigenous Lactobacillus cultures were analyzed for β-glucosidase enzyme assay in soy-drink supplemented with 2% inulin, WPC 70 and SMP respectively under optimal conditions at 24h of incubation. These cultures having β-glucosidase content produced a yellow color due to the reaction caused by releasing p-nitrophenol from the substrate pNPG as represented in Figure 2. The lactic cultures employed in the study generated various levels of β-glucosidase activity in plain soy-drink without any supplementation and also in soy-drink supplemented with inulin, WPC and SMP throughout their growth period under optimal conditions at 24 h of incubation. L. fermentum K16 showed the highest enzyme activity of 0.154 µM/mL/min in 2% inulin supplemented soy-drink, 0.122 µM/mL/min in 2% rate of WPC 70 followed by the activity of 0.0560 µM/mL/min in 2% SMP supplemented soy-drink as compared to rest of the cultures and was much higher than soy-drink without any supplementation respectively. Hence, from the univariate analysis of variance, it was derived that β-glucosidase activity in all the eight cultures was significantly highest (p≤0.05) after supplementation with inulin as compared to WPC 70 and SMP incorporation.
 |
Figure 2: β-glucosidase activity of Lactobacillus cultures in soy milk supplemented with inulin, SMP, WPC 70; Control- plain soy milk without any supplementation. |
In the study, the findings are in accordance with Donkor et al.35. Tsangalis et al.24 reported that fermented soy-drink showed different levels of β-glucosidase activity in correlation with the starter organisms applied. Hati et al.28 also reported high β-glucosidase activity in soy-drink added with WPC 70 at 2% level compared to soy-drink without any supplementation during the fermentation. In a study conducted by Hati et al.14 univariate analysis was conducted at the level = 0.05, exhibited a response to β-glucosidase activity by L. rhamnosus C6 was high (p≤0.05) as compared to other cultures proving that β-glucosidase activity is strain-dependent. With concordance to our study, Otieno et al.18 reported high β-glucosidase activity of Lactobacillus cultures viz. L. rhamnosus 4692, L. casei ASCC 290, L. acidophilus 33200, and B. animalis BB12. The β-glucosidase enzymatic potentiality relies on the strain employed and specific nutritional element provided by the addition of inulin/WPC/SMP for supporting their exponential growth and further this bioconversion could be improved at a higher margin by prebiotic (inulin) or WPC 70 supplementation in soy-drink thereby providing extra nutritional compounds to the organisms28. Deglucosylation can release attractive flavor compounds from precursors that are glucosylated, thereby increasing the content and bioavailability of plant metabolites and dietary toxins, which are health-promoters, contributing significantly to the dietary and sensory parameters of fermented food6.
β-Galactosidase Activity of Lactobacillus Cultures in Soy-Drink with Nutritional Factors
β-galactosidase enzymes regulate the transgalactosylation reactions to synthesize galactooligosaccharides from lactose catalysis found in milk. This activity exhibits a better industrial product for people suffering from lactose intolerance36. From all the indigenous Lactobacillus cultures, L. acidipiscis K27A showed the highest activity of 0.339 µM/mL/min in inulin supplemented soy-drink. Comparatively, L. rhamnosus K4E produced a high enzyme activity of 0.306 µM/mL/min in WPC 70 supplemented soy-drink, followed by an enzyme activity of 0.191 µM/mL/min by L. fermentum K7 in SMP supplemented soy-drink (Figure 3) respectively. Hence, from the univariate analysis of variance, it was derived that β-galactosidase activity in all the eight cultures was significantly highest (p≤0.05) after supplementation with inulin as compared to WPC 70 and SMP incorporation. Makwana et al.21 stated the β-galactosidase activity of L. helveticus MTCC 5463 increased significantly till 24h incubation, which was in agreement to our findings, where β-galactosidase activity was high after fermentation of 24h with absorbance at 1.44 absorbance for the Lactobacillus cultures in comparison to 48h with 0.467 O.D. and 72h with 0.193 O.D. By optimizing the lysozyme and sonication method for the liberation of protein from the cells of L. plantarum evaluating the production and characterization of β-galactosidase from it, claiming sonication to be the most prominent method37. Sangwan et al.38 reportedly applied various disciplines for disrupting cells of S. thermophilus, screened out from different dairy food products for releasing β-galactosidase and further biosynthesis of galactooligosaccharides by enzyme extraction employing whey added with various concentrations of lactose (substrate).
 |
Figure 3: β-galactosidase activity of Lactobacillus cultures in soy milk supplemented with inulin, SMP, WPC 70; Control- plain soy milk without any supplementation. |
Conclusion
The viability of all the indigenous Lactobacillus strains was significantly enhanced in soy-drink supplemented with the nutritional factors during fermentation. L. fermentum K7 showed the highest proteolytic activity (1.17 O.D.) in soy-drink supplemented with WPC 70. In comparison to the rest of the Lactobacillus cultures in the study, L. fermentum K16 showed maximum β-glucosidase enzyme activity (0.154µM/mL/min) and L. acidipiscis K27 produced the highest β-galactosidase activity (0.339 µM/mL/min) in soy-drink supplemented with inulin. Furthermore, this combination of nutritional supplements with the indigenous lactic cultures may be implemented to prepare novel fermented soy foods.
Acknowledgment
The authors acknowledge support assisted by the Department of Biotechnology (DBT) sponsored Bioinformatics Infrastructure Facility (BIF) Center, North-Eastern Hill University, Tura Campus, Tura, Meghalaya for the analyzing and interpretation of the data.
Funding Sources
The author(s) received no financial support for this doctoral research, authorship, and/or publication of this article.
Conflicts of Interest
None of the authors has any conflicts of interest to declare.
References
- Pyo, Y.H., Song, S.M. Physicochemical and sensory characteristics of a medicinal soy yogurt containing health-benefit ingredients.J Agric Food Chem. 2008; 57(1): 170-175. https://doi.org/10.1021/jf8026952.
CrossRef - Roberfroid, M. Prebiotics: the concept revisited.The J Nutr. 2007; 137(3): 830S-837S. https://doi.org/10.1093/jn/137.3.830S.
CrossRef - Guggisberg, D., Cuthbert-Steven, J., Piccinali, P., Bütikofer, U., Eberhard, P. Rheological, microstructural and sensory characterization of low-fat and whole milk set yoghurt as influenced by inulin addition. Int Dairy J.2009; 19(2): 107-115. https://doi.org/10.1016/j.idairyj.2008.07.009
CrossRef - Pham, T.T., Shah, N.P. Skim milk powder supplementation affects lactose utilization, microbial survival and biotransformation of isoflavone glycosides to isoflavone aglycones in soymilk by Lactobacillus.Food Microbiol. 2008; 25(5): 653-661. https://doi.org/10.1016/j.fm.2008.04.004.
CrossRef - Kaur, A., Bajwa, U., Goraya, R.K., Singh, A. Whey Protein Concentrate as a Substitute to Skim Milk Powder in Soy Yoghurt. Agric Res Technol. 2017; 6(4): 555691.
CrossRef - Michlmayr, H., Kneifel, W. β-Glucosidase activities of lactic acid bacteria: mechanisms, impact on fermented food and human health. FEMS Microbiol Lett. 2014; 352(1): 1-10. https://doi.org/10.1111/1574-6968.12348.
CrossRef - Vasiljevic, T., Jelen, P. Production of β-galactosidase for lactose hydrolysis in milk and dairy products using thermophilic lactic acid bacteria. Innovative Food Sci Emerging Technol. 2001; 2(2): 75-85. https://doi.org/10.1016/S1466-8564(01)00027-3.
CrossRef - Dave, R.I., Shah, N.P. Ingredient supplementation effects on viability of probiotic bacteria in yogurt.J Dairy Sci. 1998; 81(11): 2804-2816. doi:10.3168/jds.S0022-0302(98)75839-4.
CrossRef - Shihata, A. Shah, N.P. Proteolytic profiles of yogurt and probiotic bacteria. Int Dairy J.2000; 10(5-6): 401-408. https://doi.org/10.1016/S0958-6946(00)00072-8.
CrossRef - Smid, E.J., Poolman, B., Konings, W.N. Casein utilization by lactococci. Appl Environ Microbiol. 1991; 57(9): 2447.
CrossRef - Mishra, B.K., Hati, S., Das, S., Patel, K. Bio-Diversity of Lactobacillus cultures associated with the traditional ethnic fermented foods of West Garo Hills, Meghalaya, India. Int J Curr Microbiol Appl Sci. 2017; 6(2): 1090-1102. http://dx.doi.org/10.20546/ijcmas.2017.602.123.
CrossRef - Mishra, B.K., Hati, S., Das, S., Kumari, R. Evaluation of probiotic potentials of Lactobacillus isolated from traditional fermented foods of Garo Hills, Meghalaya, India. Medical Microbiol. 2018; 29(3): 120-128. DOI:10.1097/MRM.0000000000000139.
CrossRef - Mishra, B.K., Hati, S., Das, S., Prajapati, J.B. Biofunctional Attributes and Storage Study of Soy Milk Fermented by Lactobacillus rhamnosus and Lactobacillus helveticus. Food Technol. Biotechnol. 2019; 57(3): 399-407. https://doi.org/10.17113/ftb.57.03.19.6103.
CrossRef - Hati, S., Vij, S., Mandal, S., Malik, R.K., Kumari, V., Khetra, Y. α‐Galactosidase Activity and Oligosaccharides Utilization by Lactobacilli during Fermentation of Soy Milk. J Food Process Preserv. 2014; 38(3): 1065-71. doi:10.1111/jfpp.12063.
CrossRef - Indian Standards. Methods of test for dairy industry part-Irapid examination of milk, Indian Standards Institution, New Delhi (1479) (1960).
- IDF 146. Yogurt-Identification of characteristic microorganisms (Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus), International Standard, (2003).
- Donkor, O.N., Henriksson, A., Vasiljevic, T., Shah, N.P. Proteolytic activity of dairy lactic acid bacteria and probiotics as determinant of growth and in vitro angiotensin-converting enzyme inhibitory activity in fermented milk.Le Lait. 2007; 87(1): 21-38. https://doi.org/10.1051/lait:2006023.
CrossRef - Otieno, D.O., Ashton, J.F., Shah, N.P. Evaluation of enzymic potential for biotransformation of isoflavone phytoestrogen in soymilk by Bifidobacterium animalis, Lactobacillus acidophilus and Lactobacillus casei. Food Res Int. 2006; 39(4): 394-407.
CrossRef - Hati, S., Vij, S., Singh, B.P., Mandal, S. β‐Glucosidase activity and bioconversion of isoflavones during fermentation of soymilk.J Sci Food Agric. 2015; 95(1): 216-220. https://doi.org/10.1002/jsfa.6743.
CrossRef - Prasad, L.N., Ayyash, M.M., Shah, N.P. Effect of carbon and nitrogen sources on growth of Bifidobacterium animalis Bb12 and Lactobacillus delbrueckii bulgaricus ATCC 11842 and production of β-galactosidase under different culture conditions. Int Food Res J. 2011; 18: 373-380.
- Makwana, S., Hati, S., Parmar, H., Aparnathi, K.D. Process Optimization for the Production of β-Galactosidase using Potential Lactobacillus Int J Curr Microbiol Appl Sci. 2017; 6(8): 1454-1469. https://doi.org/10.20546/ijcmas.2017.608.176.
CrossRef - Farnworth, E.R., Mainville, I., Desjardins, M.P., Gardner, N., Fliss, I., Champagne, C. (2007). Growth of probiotic bacteria and bifidobacteria in a soy yogurt formulation. Int J Food Microbiol. 116(1): 174-81. https://doi.org/10.1016/j.ijfoodmicro.2006.12.015.
CrossRef - Brain, J.B. Microbiology of fermented foods. 2nd. New York, USA: Blackie Academic and Professional; 1999. 1-851.
- Tsangalis, D., Shah, N.P. Metabolism of oligosaccharides and aldehydes and production of organic acids in soymilk by probiotic bifidobacteria.Int J Food Sci Technol. 2004; 39(5): 541-554. https://doi.org/10.1111/j.1365-2621.2004.00814.x
CrossRef - Stijepić, M., Glušac, J., Djurdjević-Milošević, D., 2013. Effect of honey addition on rheological properties of probiotic soy yoghurt. Glasnik hemičara, tehnologa i ekologa Republike Srpske. 2013; 5(9).
CrossRef - Kothari, S.L. Studies on fermented soymilk products. III. Activity of cheese and yoghurt cultures in soymilk and fortified soymilk.Indian J Microbiol. 1975; 15(1): 18-26.
- Rajasekaran, M., Rajor, R.B. Supplementation of soymilk with skim milk develop youghurt like product.Asian J Dairy Res. 1989; 8(3): 155-159.
- Hati, S., Patel, N., Patel, K., Prajapati, J.B. Impact of whey protein concentrate on proteolytic lactic cultures for the production of isoflavones during fermentation of soy milk. J Food Process Preserv.2017; 41(6): e13287. https://doi.org/10.1111/jfpp.13287.
CrossRef - Subrota, H., Shilpa, V., Brij, S., Vandna, K., Surajit, M. Antioxidative activity and polyphenol content in fermented soy milk supplemented with WPC-70 by probiotic Lactobacilli. Int Food Res J., 2013; 20(5): 2125.
- Pan, X., Wu, T., Zhang, L., Cai, L., Song, Z. Influence of oligosaccharides on the growth and tolerance capacity of lactobacilli to simulated stress environment. Lett Appl Microbiol. 2009; 48(3): 362-367. https://doi.org/10.1111/j.1472-765X.2008.02539.x.
CrossRef - Gobbetti, M., De Angelis, M., Corsetti, A., Di Cagno, R. Biochemistry and physiology of sourdough lactic acid bacteria. Trends Food Sci Technol. 2005; 16(1-3): 57-69.
CrossRef - Hati, S., Vij, S., Mishra, B.K., Mandal, S., Choubey, C.S. Effect of antioxidative activity and polyphenol content in fermented soy milk. Bioved. 2012; 24: 225–234.
- Beganović, J., Kos, B., Pavunc, A.L., Uroić, K., Džidara, P., Šušković, J. Proteolytic activity of probiotic strain Lactobacillus helveticus M92. Anaerobe. 2013; 20: 58-64.
CrossRef - Kumari P., Vij, S. Growth and antimicrobial activity of proteolytic probiotic Lactobacillus rhamnosus C6 in soymilk and whey. Indian J Dairy Sci. 2015; 68: 229–238.
- Donkor, O.N., Henriksson, A., Singh, T.K., Vasiljevic, T., Shah, N.P. ACE-inhibitory activity of probiotic yoghurt. Int Dairy J. 2007: 17(11): 1321-31. https://doi.org/10.1016/j.idairyj.2007.02.009.
CrossRef - Kamran, A., Bibi, Z., Aman, A., Qader, S.A. Lactose hydrolysis approach: isolation and production of β-galactosidase from newly isolated Bacillus strain B-2. Biocatal Agric Biotechnol. 2016; 5: 99-103. https://doi.org/10.1016/j.bcab.2015.12.010.
CrossRef - Kara, F.I.R.A.T. Release and characterization of beta-galactosidase from Lactobacillus plantarum. Ankara, Turkey: The graduate school of natural and applied sciences, Middle East Technical University, PhD thesis. 2004.
- Sangwan, V., Tomar, S.K., Ali, B., Singh, R.R., Singh, A.K. Production of β-galactosidase from Streptococcus thermophilus for galactooligosaccharides synthesis. J Food Sci Technol. 2015; 52(7): 4206-4215.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








