

Introduction
Terminalia ferdinandiana (Exell, Combretaceae), is an endemic plant of Australia, with edible fruits that are extremely rich in antioxidant compounds.1-6 Like its counterparts from the genus Terminalia, this plant has a rich history of being utilised as a traditional medicine by the Australian aboriginal communities. Fruits of this plant have been used to prepare various ailments for the cure of headaches, to alleviating symptoms of colds and flu and as an antiseptic.1-8 This plant has been also used for its medicinal properties in the same way as T. carpentariae, another Australia native Terminalia species.9
Recent studies have reported on a number of bioactive properties of T. ferdinandiana which support many of the traditional medicinal claims of this plant by the Australian Aboriginal communities.10-12 Some of these reports also indicated that polar solvent extracts from T. ferdinandiana fruit were effective in inhibiting both Gram-positive and Gram-negative bacteria.10-12
Protective effects of T. ferdinandiana fruit extracts on oxidative stress and inflammatory pathways have been also reported by other authors.13-14 The potent biological activity observed in T. ferdinandiana is attributed to the presence of enhanced levels of antioxidant bioactive compounds.15-16 Some of the bioactive phytochemicals detected in T. ferdinandiana fruit and leaves include ascorbic acid, ellagic acid, gallic acid, α-tocopherol, ethyl gallate, chebulic acid, corilagin, hydroxycinnamic acid, lutein, tannins, chebulagic acid, exifone, punicalin, castalagin, appanone A-7 methyl ether, xanthotoxin, phthalane, saponins, flavonoids, and terpenes.1-19 Elevated antioxidant activity was confirmed by early studies by Netzel and collaborators (2007) who measured the antiradical properties by the TROLOX Equivalent Antioxidant Capacity (TEAC) assay.15 The antioxidant capacity of 567 T ferdinandiana fruit from 45 geographic sites was also reported demonstrating the important antioxidant activity of this plant.2, 20-21 Recently, TPC of methanolic extracts of T. ferdinandiana fruits and leaf (obtained by accelerated solvent extraction) have been reported to be 12.2 and 11.7 g GAE/100 g DW.22
One of the most prominent antioxidant phytochemicals present in T. ferdinandiana is ascorbic acid or vitamin C, which is essential for human health. Due to its strong antioxidant properties, ascorbic acid neutralizes reactive oxygen species, prevents the generation of new free radicals by suppressing relevant molecular pathway and assists in the recycling of other antioxidants.20-21 Vitamin C also plays an important immunomodulatory functions such as regulation of macrophage activity, reduction of inflammatory mediators, and imparting direct bacteriostatic effect at high concentrations.22
High levels of ascorbic acid (>14% DW) have been reported in T. ferdinandiana fruits.5-6 Other studies have also reported a wide range of ascorbic acid levels in T. ferdinandiana fruits (3.5 – 5.9% FW) and in the range of 0.1 – 5.3% FW.1-4, 5,23 Levels of ascorbic acid in the T. ferdinandiana fruit were observed at significantly higher concentrations than other well-known natural sources of ascorbic acid like citrus fruit (0.5% FW ascorbic acid)24, Acerola fruit (1.0 and 1.4% FW)24-25 and Camu-Camu (1.8% FW).24-25
As the consumer awareness on the health promoting activities of the T. ferdinandiana fruit and its products, increasing demands for new applications are driving food industry to find novel applications of T. ferdinandiana as a functional food ingredient. Currently, only frozen puree and freeze-dried powder of T. ferdinandiana fruit are commercially available; however, anecdotal evidences and recent studies have indicated that other tissues such as leaves, seed coats and kernels could also be used for food and other applications.22, 26 The leaves of T. ferdinandiana could be a great candidate for such novel applications. Anecdotally, there are accounts of using broken T. ferdinandiana leaves for cleaning by scrubbing hands within the Indigenous Australians indicating potential antimicrobial efficacy of the leaves. A recent study, evaluated the antimicrobial activity of extracts were prepared from different T. ferdinandiana tissues including fruits, leaves, seedcoats, and barks. It was observed that both fruit and leaf extract exhibited superior antimicrobial activity, against common foodborne bacteria.22, 26
As T. ferdinandiana is a semi-deciduous tree, it drops its leaves in the dry season, and spouts new leaves at the beginning of the wet season. The characteristic morphology of the leaves should make them easier to harvest than fruit. Leaves of Terminalia plants are usually spirally organised (often crowded) at the ends of the branches, sometimes on short shoots. In fact, the genus name ‘Terminalia’ comes from the Latin word ‘Terminus’ relating to the fact that the leaves are located at the very tips of the shoots. Mature T. ferdinandiana leaves are usually large in size, making it possible to collect larger volume of raw material. Although several studies have shown the bioactive potentials of T. ferdinandiana leaves, information on the effect of maturity of leaves on their bioactive compounds or bioactivity is not available.
The objective of this work was to provide with information on morphology, antimicrobial activity, total phenolic and ascorbic acid content in T. ferdinandiana leaves over different stages of maturity.
Materials and Methods
Plant Material
Fieldwork for this study was conducted in undeveloped bushland in the northwestern corner of the Charles Darwin University, Casuarina campus (Darwin, Northern Territory, Australia) (see Figure 1). The study site is within one kilometre of the coastal shoreline, comprising open savanna woodland with incipient monsoon forest along old streamlines and is representative of T. ferdinandiana habitat.23 The mean annual rainfall from 1995 to 2020 was 1768 mm and mean daily temperatures ranged from a minimum of 23.2 to a maximum of 32.1°C (Bureau of Meteorology 2020).
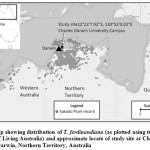 |
Figure 1: Map showing distribution of T. ferdinandiana (as plotted using tree records from Atlas of Living Australia) and approximate locate of study site at Charles Darwin University, Darwin, Northern Territory, Australia. |
Leaf samples were harvested from November 2017 to June 2018. A total of 15 different maturity stages of leaves (10 leaves per stage) were collected from 10 individual trees (AT1 to AT10). Stage 1 to 4 contain immature leaves; stage 5 to 10 contain mature leaves and stage 11 to 15 contain senescing leaves. Leaves of stages 12 to 15 were collected from fewer than 10 individual trees as some of the trees had dropped their leaves ahead of others. Details of the harvesting time are provided in Table 1. For the current study, 5 maturity stages, were selected from 3 individual trees (10 leaves per stage per tree) for analysis. The selected maturity stages for the current study allowed us to look at leaves at immature stage (e.g. stages 2 and 3), mature stage (e.g. stage 6 and 10) and senescing stage (e.g. stage 15) (see Figure 2).
 |
Figure 2: The selected maturity stages of the current study. Stages 2 and 3 represent the immature leaves, stages 6 and 10 represent the mature leaves and stage 15 represent senescing leaves.Click here to View Figure |
Table 1: Time of Collection of Terminalia Ferdinandiana Leaf Samples.
| Maturity stages | Time of collection | |
| Immature leaves* | 1 | 14 Nov 2017 |
| 2 | 14 Nov 2017 | |
| 3 | 14 Nov 2017 | |
| 4 | 14 Nov 2017 | |
| Mature leaves | 5 | 9 Jan 2018 |
| 6 | 14 Feb 2018 | |
| 7 | 12 Mar 2018 | |
| 8 | 14 Apr 2018 | |
| 9 | 14 May 2018 | |
| 10 | 21 May 2018 | |
| Senescing leaves | 11 | 30 May 2018 |
| 12** | 6 Jun 2018 | |
| 13*** | 13 Jun 2018 | |
| 14*** | 21 Jun 2018 | |
| 15*** | 26 Jun 2018 |
*These samples were collected on the same time point as the newly sprouted leaves of stage 1. Leaves were divided into stage 2, 3 and 4 on the increase in sizes.
**Leaves from stage 12 were collected from 6 individual trees (10 leaves per tree).
***Leaves from stage 13, 14 and 15 were collected from 4 individual trees (10 leaves per tree).
Fieldwork for this study was conducted in undeveloped bushland in the northwestern corner of the Charles Darwin University, Casuarina campus (Darwin, Northern Territory, Australia) (see Figure 1). The study site is within one kilometre of the coastal shoreline, comprising open savanna woodland with incipient monsoon forest along old streamlines and is representative of T. ferdinandiana habitat.23 The mean annual rainfall from 1995 to 2020 was 1768 mm and mean daily temperatures ranged from a minimum of 23.2 to a maximum of 32.1°C (Bureau of Meteorology 2020).
Leaf samples were harvested from November 2017 to June 2018. A total of 15 different maturity stages of leaves (10 leaves per stage) were collected from 10 individual trees (AT1 to AT10). Stage 1 to 4 contain immature leaves; stage 5 to 10 contain mature leaves and stage 11 to 15 contain senescing leaves. Leaves of stages 12 to 15 were collected from fewer than 10 individual trees as some of the trees had dropped their leaves ahead of others. Details of the harvesting time are provided in Table 1. For the current study, 5 maturity stages, were selected from 3 individual trees (10 leaves per stage per tree) for analysis. The selected maturity stages for the current study allowed us to look at leaves at immature stage (e.g. stages 2 and 3), mature stage (e.g. stage 6 and 10) and senescing stage (e.g. stage 15) (see Figure 2).
Morphological Parameters
Length and width of 10 individual leaves of each maturity stage from three individual trees were measured using a digital calliper (Craftright Engineering Works, Jiangsu, China) followed by measuring the weight on laboratory scales (Sartorius CP224S analytical balance with readability precision 0.01g , Gottingen, Germany). The leaves were frozen at -80 ºC and then freeze dried at -50 ºC for 48h (CSK Climatek, Darra, QLD, Australia). After freeze drying, around 2 g of dry leaves were ground using a Retsch MM400 ball mill (Retsch GmbH, Haan, Germany) at a speed of 30 Hz for 1 min.
Extraction of Bioactive Constituents
Bioactive constituents from T. ferdinandiana leaves were extracted by either methanol or deionized water. Triplicate samples (approx. 0.5 g) of the freeze-dried powders were accurately weighed into separate centrifuge tubes and individually blended with 5 ml of each solvent. After 30 seconds of vortex-mixing, the tubes were sonicated in an ultrasonic bath (Elma Schmidbauer GmbH, Ruiselede, Belgium) for 5 min at room temperature, followed by another 5 min of gentle agitation. The slurry was subsequently centrifuged at 3900 rpm for 5 min using an Eppendorf 5180 centrifuge (Eppendorf, Hamburg, Germany). The supernatant was carefully transferred and collected while the residues extracted twice again. After 3 times of extractions, a total volume of 15 ml of the crude extracts were combined and evaporated in a rotatory evaporator (Genevac Ltd, Ipswich, Suffolk, England) at 40°C. The dried extracts were freshly reconstituted in 5 mL of aqueous 20% v/v methanol (for methanol extracts) or deionized water (for water extracts) prior to analysis of antimicrobial activities. The reconstitute extract was stored at biomedical freezer (MDF U5312, PHCbi, Panasonic) and daily monitored using a digital thermometer.
Antimicrobial Screening
A total of three pathogenic microbial strains were tested in this study to evaluate the antimicrobial activity of T. ferdinandiana leaf extracts: Staphylococcus aureus NTCC 6571, a Gram-positive bacteria; Escherichia coli NTCC 9001, a Gram-negative bacteria and Candida albicans ATCC 90028 a fungi. The bacterial strains were purchased from American Type Culture Collection, USA or National Collection of Type Cultures (NCTC), UK. Well diffusion assay was used to evaluate the antimicrobial activity of the leaf extracts followed the method published by Phan and collaborators (2019).27 The inhibition zone was measured using a 150 mm Digital Calliper (Craftright Engineering Works, Jiangsu, China). MIC will be suggested for further investigations uing a widen ranges of microorganisms. All plates were incubated overnight in triplicate (see Supplementary material S2).
Supplementary 2: Representative photos show (B) the inhibition of methanol extracts and (A) antibiotic solution (10 μl of Penicillin and streptomycin at 1g each/10 mL methanol) against Staphyloccocus areus and negative control 20% methanol.
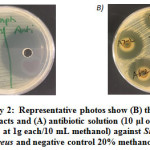 |
Supplementary 2: Representative photos show (B) the inhibition of methanol extracts and (A) antibiotic solution (10 μl of Penicillin and streptomycin at 1g each/10 mL methanol) against Staphyloccocus areus and negative control 20% methanol. |
Total Phenolic Content (TPC)
The total phenolic content (TPC) of the samples was determined using the Folin Ciocalteau (FC) method using a micro-plate reader (Sunrise Tecan, Maennedorf, Switzerland).28 Gallic acid standards ranging from 21 to 105 mg/L were prepared to establish the standard curve for quantification of TPC in the extract. TPC was expressed as mg gallic acid equivalents per gram of sample in dry weight (mg GAE/g DW).
Extraction and analysis of Ascorbic Acid
Measurement of vitamin C content in T. ferdinandiana leaf extracts was conducted by utilizing ultra-high-performance liquid chromatographic–photodiode array (UHPLC-PDA) methodology.29-30 (See chromatogram S1).
Supplementary 1: UHPLC-PDA acquired at 245 nm of (A) ascorbic acid standard and (B) the leaf extract
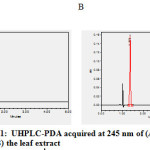 |
Supplementary 1: UHPLC-PDA acquired at 245 nm of (A) ascorbic acid standard and (B) the leaf extract. |
Data Analysis
Descriptive statistics (average, minimum and maximum values, and standard deviation), principal component analysis (PCA) and multiple linear regression (MLR) were applied to inspect the relevant and interpretable structure in the data set associated with the variables measured in the leaf samples at the different maturities. The optimum number of components in both PCA and MLR analysis was determined internal cross validation (leave one out).31 The Unscrambler software (version 10.5, CAMO, Norway) was used to develop the PCA and MLR regression models. Samples were standardised using the pre-processing 1/SD.31
Results and discussion
The current study analysed T. ferdinandiana leaves at 5 maturity stages namely immature leaves at stages 2 and 3; mature leaves at stages 6 and 10, and senescing leaves at stage 15. Leaves from stages 1 to 4 were collected at the same time as the freshly sprouted leaves (e.g. stage 1). These immature leaves were then divided into 4 stages according to their sizes. From stages 5 to 15, leaves were collected at different time points and grouped according to their stage of development/maturity (Table 1).
Morphological analysis showed that at maturity stage 2, irrespective of the tree, leaves had the smallest length and width, both of which showed a gradual increase in the leaves of stage 3. However, leaves at maturity stages 6, 10 and 15 did not have a clear trend and showed high variability in length, width and weight (Table 2). Tree-to-tree variability has been also observed in T. ferdinandiana fruits. The high degree of variability observed in the leaves and fruits could be attributed to the fact T. ferdinandiana is a wild harvested plant and often undergoes wide cross-pollination. Interestingly, the variability of the morphological parameters seemed to decrease with the maturity stage of the leaves which is indicated by a lower coefficient of variation (% CV) in the morphological parameters measured (Table 5).
Table 2: Morphological data for Terminalia ferdinandiana leaves at various stages of maturity.
| Tree ID | Maturity stage | Length (mm) | Width (mm) | Weight (g) |
| AT2 | 2 | 58.7 ± 6.5 | 36.8 ± 5.1 | 0.4 ± 0.1 |
| 3 | 112.5 ± 8.6 | 95.5 ± 10.9 | 1.9 ± 0.3 | |
| 6 | 159.6 ± 16.5 | 123.4 ± 9.6 | 4.0 ± 0.9 | |
| 10 | 149.5 ± 18.0 | 122.6 ± 15.8 | 3.4 ± 0.6 | |
| 15 | 124.8 ± 30.0 | 106.3 ± 16.4 | 2.4 ± 0.9 | |
| AT5 | 2 | 80.5 ± 7.7 | 50.9 ± 7.1 | 0.8 ± 0.2 |
| 3 | 133.4 ± 20.4 | 98.9 ± 18.1 | 2.5 ± 0.8 | |
| 6 | 184.6 ± 17.1 | 126.3 ± 16.1 | 5.1 ± 0.8 | |
| 10 | 162.4 ± 24.1 | 118.1 ± 27.7 | 3.6 ± 1.1 | |
| 15 | 189.4 ± 28.4 | 123.9 ± 17.1 | 4.1 ± 1.1 | |
| AT9 | 2 | 73 ± 19.9 | 46.2 ± 12.2 | 1.2 ± 0.8 |
| 3 | 138.1± 25.5 | 91.4 ± 15.8 | 2.5 ± 0.9 | |
| 6 | 187 ± 23.8 | 136.1 ± 14.8 | 5.0 ± 1.0 | |
| 10 | 168.9 ± 48.4 | 119.7 ± 21.5 | 3.6 ± 1.7 | |
| 15 | 135.3 ± 17.7 | 109.9 ± 19.0 | 3.6 ± 1.3 |
Results are mean ± SD (n = 10)
AT indicate the tree ID of the individual trees collected from the study site at Charles Darwin University, Darwin, Northern Territory, Australia.
The ascorbic acid levels determined in leaves at maturity stage 6, 10 and 15 were between the ranges of 22.5 to 34.8 mg/100 g DW (Table 3) whereas no peak of ascorbic acid was observed in the UHPLC-PDA chromatogram for leaves sourced from stages 2 and 3, respectively. This data suggested that levels of ascorbic acid in these immature leaves might be present either at a relatively low concentration or lower than the limit of detection (LOD = 0.1 parts per million) of the method used for the analysis. The lower levels of ascorbic acid in T. ferdinandiana leaves compared to fruits have been previously reported by other authors.5-6 It is well known that ascorbic acid is important bioactive compound having important roles in the plant (e.g. redox reactions, cofactor of enzymes, photosynthesis, hormone biosynthesis, antioxidant function).32-35 Variation in the ascorbic acid content has been reported among different tissues and organs in the same tree by other authors. 32-35 High concentration of ascorbic acid was found in tissues such as leaves and flowers compared with those photosynthetically active such as stems and roots where higher concentrations are present in the meristematic tissues and reproductive organs (e.g. flowers, young fruits). 32-35 It has been also reported that the concentration of ascorbic acid might be affected by the environment and developmental stages of the organ (i.e. mature vs immature fruits).36 These factors might explain the observed variations in ascorbic acid in the leaf samples analysed.
Table 3: Total ascorbic acid and total polyphenol content Terminalia ferdinandiana leaves at various stages of maturity.
| Tree ID | Maturity stage | Total ascorbic acid (mg/100 g DW) | Total polyphenol content (mg GAE/g DW) | |
| Methanolic extracts | Water extracts | |||
| AT2 | 2 | ND | 533.4 ± 1.5 | 263.1 ± 0.8 |
| 3 | ND | 598.6 ± 1.4 | 301.9 ± 1.3 | |
| 6 | 23.0 ± 0.1 | 279.8 ± 2.6 | 253.9 ± 2.2 | |
| 10 | 29.6 ± 0.5 | 384.9 ± 2.4 | 274.9 ± 0.5 | |
| 15 | 34.8 ± 0.3 | 407.5 ± 1.9 | 295.9 ± 0.6 | |
| AT5 | 2 | ND | 294.8 ± 1.5 | 222.2 ± 0.5 |
| 3 | ND | 330.3 ± 0.8 | 231.8 ± 0.6 | |
| 6 | 32.3 ± 0.8 | 237.3 ± 1.6 | 210.3 ± 2.1 | |
| 10 | 34.9 ± 0.3 | 283.9 ± 0.9 | 220.3 ± 2.5 | |
| 15 | 35.5 ± 0.1 | 247.5 ± 2.1 | 215.6 ± 0.4 | |
| AT9 | 2 | ND | 388.7 ± 2.3 | 276.2 ± 0.3 |
| 3 | ND | 579.6 ± 0.3 | 319.6 ± 0.2 | |
| 6 | 22.5 ± 0.4 | 359.8 ±1.0 | 258.4 ± 0.2 | |
| 10 | 32.6 ± 0.5 | 323.8 ±0.9 | 259.0 ± 0.4 | |
| 15 | 34.7 ± 0.7 | 379.9 ± 0.7 | 283.2 ± 0.5 | |
The total phenolic content (TPC) of the leaf extracts ranged from 210.3 to 598.6 mg/g DW. The TPC values varied from 237.3 to 598.6 mg GAE/g DW in the methanolic extracts and from 210.3 to 319.6 mg GAE/g DW in the water extracts. TPC was higher in the leaf methanolic extracts compared to the water extracts. Similar results were reported by Akter and collaborators (2019). TPC showed a gradual decrease with the advance of maturity, with the highest levels observed in leaves at stage 3 irrespective of the tree in both the methanolic and water extracts (Table 3).
The antimicrobial analysis showed that the methanolic and water extracts of T. ferdinandiana leaves have strong inhibitory efficacy (e.g. inhibition zone > 13 mm)37 against the Gram-positive S. aureus , but no inhibition against the Gram-negative E. coli and the fungi C. albicans (Table 4). The antimicrobial efficacy in terms of inhibitory zones were in the range of 23.6 to 29.1 mm and 21.2 to 26.7 mm for methanolic and water extracts, respectively. A recent study has also reported that methanolic and aqueous leaf extracts of T. ferdinandiana were good inhibitors of the Gram positive Bacillus anthracis, the etiological agent of anthrax.19 Akter and collaborators (2019) observed that the methanolic and water extracts of T. ferdinandiana leaves inhibited both Gram positive and negative bacteria.26 The observed difference in the antimicrobial efficacy of T. ferdinandiana leaf extracts might be associated with the difference in the type of bioactive compounds extracted and their extent of release from the sample matrix as result of extraction method used and geographical location of the studied plant material.38-39
Table 4: Inhibition zones (mm) of methanol and water extracts against S. aureus.
| Tree ID | Maturity stage | Inhibition zones (mm) | ||
| Methanolic extracts | Water extracts | |||
| AT2 | 2 | 23.9 ± 0.8 | 21.8 ± 0.3 | |
| 3 | 27.5 ± 0.5 | 24.9 ± 0.9 | ||
| 6 | 24.9 ± 0.9 | 23.1 ± 0.9 | ||
| 10 | 26.3 ± 0.3 | 24.2 ± 0.6 | ||
| 15 | 27.2 ± 0.6 | 25.5 ± 0.4 | ||
| AT5 | 2 | 23.6 ± 1.5 | 21.8 ± 1.3 | |
| 3 | 29.1 ± 0.4 | 23.3 ± 0.6 | ||
| 6 | 24.4 ± 0.4 | 22.6 ± 0.7 | ||
| 10 | 26.1 ± 0.1 | 23.8 ± 0.4 | ||
| 15 | 27.7 ± 0.2 | 25.3 ± 0.4 | ||
| AT9 | 2 | 24.7 ± 0.4 | 21.2 ± 0.7 | |
| 3 | 28.3 ± 1.0 | 24.9 ± 0.8 | ||
| 6 | 25.7 ± 0.4 | 23.6 ± 0.7 | ||
| 10 | 26.6 ± 0.9 | 24.2 ± 0.7 | ||
| 15 | 27.2 ± 0.7 | 26.7 ± 0.9 | ||
| Antibacterial control* | 53.3 ± 0.2 | 52.8 ± 0.31 | ||
| Solvent control** | – | – | ||
Results are mean ± SD (n = 3)
*1 µg penicillin and streptomycin was used as antibacterial control.
**Solvent controls included 20% (v/v) aqueous methanol for methanolic extracts and water for water extracts.
Pearson correlation showed strong positive correlation between levels of ascorbic acid and antimicrobial inhibition exhibited by methanolic (Pearson r = 0.60) and water (Pearson r = 0.55) extracts (Table 5). This relationship indicated that ascorbic acid might be a contributing factor, in the observed antimicrobial activity of T. ferdinandiana leaf extracts. In fact, a number of studies have reported the antimicrobial efficacy of ascorbic acid against a number of pathogenic bacteria including S. aureus , E coli, Helicobacter pylori, Campylobacter jejuni and Mycobacterium tuberculosis.40-42
Table 5: Descriptive statistics of the variables measured in the leaf samples at different maturity stages
| Mean | SD | CV (%) | |
| Length (mm) | 137.2 | 45.8 | 33.3 |
| Width (mm) | 100.4 | 34.1 | 34 |
| Weight (g) | 2.9 | 1.6 | 55 |
| Total ascorbic (mg/100 g dry weight) | 31.1 | 4.8 | 15.5 |
| TPC methanolic extracts (mg GAE/g DW) | 375 | 11 | 2.9 |
| TPC water extracts (mg GAE/g DW) | 259 | 3.2 | 1.2 |
| Inhibition methanolic extracts | 26.2 | 1.6 | 6 |
| Inhibition water extracts | 23.7 | 1.4 | 6 |
The TPC of methanolic and water extracts were negatively correlated to total ascorbic acid content (Pearson r = -0.047 and -0.11 respectively). This could be explained by the fact that ascorbic acid may not be the primary contributor to TPC of T. ferdinandiana.43 As mentioned before, T. ferdinandiana fruits and leaves are also rich sources of bioactive antioxidant compounds other than ascorbic acid, such as ellagic acid, gallic acid and α-tocopherol.1-6 Significant positive correlation was observed between the morphological parameters, TPC and antimicrobial inhibition, indicating that with maturity the level of phenolics and antimicrobial efficacy increases (Table 5).
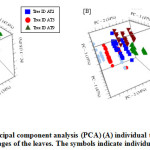 |
Figure 3: Principal component analysis (PCA) (A) individual trees and (B) maturity stages of the leaves. The symbols indicate individual subjects. |
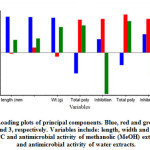 |
Figure 4: Loading plots of principal components. Blue, red and green indicate PC-1, 2 and 3, respectively. Variables include: length, width and weight of leaves, TPC and antimicrobial activity of methanolic (MeOH) extracts, TPC and antimicrobial activity of water extracts. |
The data was also analysed by principal component analysis (PCA). This method transforms a group of highly correlated variables into new data sets called principal components (PC). Then, data was interpreted using the scores and loadings. This algorithm of PCA reduces the dimensionality of the data but retains most of the variation in the data set, which increases interpretability simultaneously minimizing loss of information.31 It was observed a variation between samples obtained from different trees (Figure 3A) and maturity stages (Figure 3B), indicating the existence of natural variability between the trees. The PCA loadings allowed to interpret which variables might influence the separation between leaves (Figure 4). Most of the differences observed between the samples were found to be explained by the morphological parameters, level of ascorbic acid and antimicrobial efficacy. Furthermore, loadings in PC1 explained the differences in maturity stages where high and positive loadings corresponded well with all chemical variables such as ascorbic acid and TPC.
Multiple linear regression (MLR) analysis indicated that 97% of the variance in maturity of T. ferdinandiana leaves could be explained by the model reported in Figure 5A. The regression coefficients showed that ascorbic acid had a large influence in explaining maturity stages of the leaves, followed by both antimicrobial efficacy and TPC of the water extracts (Figure 5B). This indicates that in addition to the presence of bioactive compounds (e.g. ascorbic acid) other parameters such as antimicrobial activity and TPC might also vary with leaf maturity. Therefore, the concentration of ascorbic acid and TPC might be used as biomarkers to monitor leaf maturity in T. ferdinandiana.
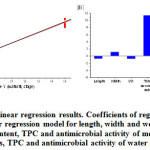 |
Figure 5: Multiple linear regression results. Coefficients of regression derived from the multiple linear regression model for length, width and weight of leaves, total ascorbic acid content, TPC and antimicrobial activity of methanolic (MeOH) extracts, TPC and antimicrobial activity of water extracts.Click here to View Figure |
Conclusion
For the first time, information on morphology, antimicrobial activity, total phenolic and ascorbic acid content in Terminalia ferdinandiana leaves was reported. Variation in the morphological parameters, TPC and ascorbic acid content with advancement of maturity was observed in the samples analysed. Both PCA and MLR analyses indicated that effect of individual trees and maturity stages, where the concentration of ascorbic acid explains the variability in maturity among leaves. However, the data in this study was not based on a large number of biological samples, and hence, is not sufficient to describe the effect of maturity on nutritional composition of T. ferdinandiana leaves. The use of statistical techniques such as PCA and MLR regression, allowed us to obtain additional information from the data set allowing for a better interpretation of the differences associated with maturity. Results from this study also indicated the pronounced inhibitory effect of T. ferdinandiana against S. aureus. Currently only the T. ferdinandiana fruit (as a freeze-dried powder and puree) is commercially available as a functional food ingredient, whereas, leaves or any other tissues, are not used for any industry applications. Leaf extracts showed promise as antimicrobial agent, suggesting that might be used as alternative to synthetic antimicrobial agents. Further studies will be recommended on T. ferdinandiana leaves, adding a large number of biological samples, trees and maturities, bioactive compounds, and anti-nutritional compounds.
Acknowledgments
The authors acknowledge the Larrakia People who are the Traditional Owners of the lands on which the Terminalia ferdinandiana was harvested, and respect the knowledge and experience the Traditional Owners hold regarding the care, harvest and use of these plants.
Conflict of Interest
The authors declare that they have no conflict of interest.
Funding
This work was supported by the Cooperative Research Centre for Developing Northern Australia (CRCNA) project – Improving the efficiency of Kakadu plum value chains to grow a robust and sustainable industry [grant number AT.2.1718031] and the Australian Research Council (ARC) Industrial Transformation Training Centre (ITTC) for Uniquely Australian Foods [Grant number: IC180100045].
References
- Konczak, I., (2009). Health benefits of Australian native foods: an evaluation of health-enhancing compounds. Rural Industries Research and Development Corporation.
- Konczak, I., Maillot, F., Dalar, A., (2014). Phytochemical divergence in 45 accessions of Terminalia ferdinandiana (Kakadu plum). Food Chemistry 151(0), 248-256.
CrossRef - Konczak, I., Zabaras, D., Dunstan, M., Aguas, P., (2010). Antioxidant capacity and hydrophilic phytochemicals in commercially grown native Australian fruits. Food Chemistry 123(4), 1048-1054.
CrossRef - Konczak, I., Zabaras, D., Dunstan, M., Aguas, P., Roulfe, P., Pavan, A., (2009). Health benefits of Australian native foods: an evaluation of health-enhancing compounds. RIRDC, Canberra, Australia.
- Williams, D.J., Edwards, D., Pun, S., Chaliha, M., Burren, B., Tinggi, U., Sultanbawa, Y., (2016). Organic acids in Kakadu plum (Terminalia ferdinandiana): The good (ellagic), the bad (oxalic) and the uncertain (ascorbic). Food Research International 89, 237-244.
CrossRef - Williams, D.J., Edwards, D., Pun, S., Chaliha, M., Sultanbawa, Y., (2014). Profiling ellagic acid content: The importance of form and ascorbic acid levels. Food Research International 66, 100-106.
CrossRef - Lim, T.K., (2012). Terminalia ferdinandiana, Edible Medicinal And Non-Medicinal Plants. Springer Netherlands, pp. 158-160.
CrossRef - Wiynjorrotj, P., Flora, S., Brown, N.D., Jatbula, P., Galmur, J., Katherine, M., Merlan, F., Wightman, G., (2005). Jawoyn Plants and Animals. Jawoyn Association, Northern Territory Government, Northern Territory, Australia.
- Conservation Commission of the Northern Territory, (1993). Traditional Aboriginal medicines in the Northern Territory of Australia conservation Commission of the Northern Territory of Australia, Darwin.
- Cock, I., Mohanty, S., (2011). Evaluation of the antibacterial activity and toxicity of Terminalia ferdinandia fruit extracts. Pharmacognosy Journal 3(20), 72-79.
CrossRef - Sirdaarta, J., Matthews, B., Cock, I.E., (2015a). Kakadu plum fruit extracts inhibit growth of the bacterial triggers of rheumatoid arthritis: Identification of stilbene and tannin components. Journal of Functional Foods 17.
CrossRef - Sirdaarta, J., Matthews, B., White, A., Cock, I.E., (2015b). GC-MS and LC-MS analysis of Kakadu plum fruit extracts displaying inhibitory activity against microbial triggers of multiple sclerosis. Pharmacognosy Communications 5.2, 100-115.
CrossRef - Chaliha, M., Sultanbawa, Y., (2019). Terminalia ferdinandiana, a traditional medicinal plant of Australia, alleviates hydrogen peroxide induced oxidative stress and inflammation, in vitro. Journal of Complementary and Integrative Medicine.
CrossRef - Tan, A.C., Hou, D.-X., Konczak, I., Tanigawa, S., Ramzan, I., Sze, D.M.Y., (2011). Native Australian fruit polyphenols inhibit COX-2 and iNOS expression in LPS-activated murine macrophages. Food Research International 44(7), 2362-2367.
CrossRef - Netzel, M., Netzel, G., Tian, Q., Schwartz, S., Konczak, I., (2007). Native Australian fruits — a novel source of antioxidants for food. Innov Food Sci Emerg 8(3), 339-346.
CrossRef - Zafrilla, P., Ferreres, F., Tomas-Barberan, F.A., (2001). Effect of processing and storage on the antioxidant ellagic acid derivatives and flavonoids of red raspberry (Rubus idaeus) jams. Journal of Agricultural and Food Chemistry 49(8), 3651-3655.
CrossRef - Courtney, R., Shalom, J., Matthews, B., Cock, I., (2015). Tannin components and inhibitory activity of Kakadu plum leaf extracts against microbial triggers of autoimmune inflammatory diseases. Pharmacognosy Journal 7, 18-31.
CrossRef - Wang, H., Cao, G., Prior, R.L., (1996). Total antioxidant capacity of fruits. Journal of Agricultural and Food Chemistry 44(3), 701-705.
CrossRef - Wright, M.H., Sirdaarta, J., White, A., Greene, C.A., Cock, I.E., (2017). GC-MS headspace analysis of Terminalia ferdinandiana fruit and leaf extracts which inhibit Bacillus anthracis growth. Pharmacognosy Journal 9(1), 73-82.
CrossRef - Wu, F., Tyml, K., Wilson, J.X., (2008). iNOS expression requires NADPH oxidase-dependent redox signaling in microvascular endothelial cells. Journal of Cellular Physiology 217(1), 207-214.
CrossRef - Wu, X., Beecher, G.R., Holden, J.M., Haytowitz, D.B., Gebhardt, S.E., Prior, R.L., (2004). Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. Journal of Agricultural and Food Chemistry 52(12), 4026-4037.
CrossRef - Akter, S., Netzel, M., Tinggi, U., Osborne, S., Fletcher, M., Sultanbawa, Y., (2019). Antioxidant Rich Extracts of Terminalia ferdinandiana Inhibit the Growth of Foodborne Bacteria. Foods 8, 281.
CrossRef - Woods, B.A., (1995). A study of the intra-specific variations and commercial potential of Terminalia ferdinandiana (the Kakadu plum). Northern Territory University, Northern Territory, Australia.
- Mezadri, T., Villaño, D., Fernández-Pachón, M.S., García-Parrilla, M.C., Troncoso, A.M., (2008). Antioxidant compounds and antioxidant activity in acerola (Malpighia emarginata DC.) fruits and derivatives. J. Food Comp. Anal. 21(4), 282-290.
CrossRef - Rufino, M.d.S.M., Pérez‐Jiménez, J., Tabernero, M., Alves, R.E., De Brito, E.S., Saura‐Calixto, F., (2010). Acerola and cashew apple as sources of antioxidants and dietary fibre. International Journal of Food Science & Technology 45(11), 2227-2233.
CrossRef - Akter, S., Netzel, M.E., Fletcher, M.T., Tinggi, U., Sultanbawa, Y. (2018). Chemical and nutritional composition of Terminalia ferdinandiana (kakadu plum) kernels: a novel nutrition source. Foods 7(60), 11.
CrossRef - Phan, A.D.T., Chaliha, M., Sultanbawa, Y., Netzel, M.E., (2019). Nutritional Characteristics and Antimicrobial Activity of Australian Grown Feijoa (Acca sellowiana). Foods 8, 376.
CrossRef - Singleton, V.L., Rossi, J.A., (1965). Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. American Journal of Enology and Viticulture 16(3), 144-158.
- Campos, F.M., Ribeiro, S.M.R., Della Lucia, C.M., Pinheiro-Sant’Ana, H.M., Stringheta, P.C., (2009). Optimization of methodology to analyze ascorbic and dehydroascorbic acid in vegetables. Quimica Nova 32, 87-91.
CrossRef - Spinola, V., Mendes, B., Camara, J.S., Castilho, P.C., (2012). An improved and fast UHPLC-PDA methodology for determination of L-ascorbic and dehydroascorbic acids in fruits and vegetables. Evaluation of degradation rate during storage. Analytical and Bioanalytical Chemistry 403(4), 1049-1058.
CrossRef - Cozzolino, D., Power, A., Chapman, J., (2019). Interpreting and Reporting Principal Component Analysis in Food Science Analysis and Beyond. Food Analytical Methods 12(11), 2469-2473.
CrossRef - Valpuesta, V., Botella, M.A., (2004). Biosynthesis of L-ascorbic acid in plants: new pathways for an old antioxidant. Trends in Plant Science 9(12), 573-577.
CrossRef - Smirnoff, N., Pallanca, J.E., (1996). Ascorbate metabolism in relation to oxidative stress. Biochemical Society Transactions 24(2), 472-478.
CrossRef - Smirnoff, N., Wheeler, G.L., (2000). Ascorbic Acid in Plants: Biosynthesis and Function. Critical Reviews in Plant Sciences 19(4), 267-290.
CrossRef - Smirnoff, N., (2011). Chapter 4 – Vitamin C: The Metabolism and Functions of Ascorbic Acid in Plants, in: Rébeillé, F., Douce, R. (Eds.), Advances in Botanical Research. Academic Press, pp. 107-177.
CrossRef - Dumas, Y., Dadomo, M., Di Lucca, G., Grolier, P., (2003). Effects of environmental factors and agricultural techniques on antioxidant content of tomatoes. Journal of the Science of Food and Agriculture 83(5), 369-382.
CrossRef - Mosbah, H., Louati, H., Boujbiha, M.A., Chahdoura, H., Snoussi, M., Flamini, G., Ascrizzi, R., Bouslema, A., Achour, L., Selmi, B., (2018). Phytochemical characterization, antioxidant, antimicrobial and pharmacological activities of Feijoa sellowiana leaves growing in Tunisia. Industrial Crops and Products 112, 521-531.
CrossRef - Trombetta, D., Castelli, F., Sarpietro, M.G., Venuti, V., Cristani, M., Daniele, C., Saija, A., Mazzanti, G., Bisignano, G., (2005). Mechanisms of antibacterial action of three monoterpenes. Antimicrobial Agents and Chemotherapy 49(6), 2474-2478.
CrossRef - Koohsari, H., Ghaemi, E.A., Sadegh Sheshpoli, M., Jahedi, M., Zahiri, M., (2015). The investigation of antibacterial activity of selected native plants from North of Iran. Journal of medicine and life 8(Spec Iss 2), 38-42.
- Isela, S., Sergio, N., Jose, M., Rene, H., Claudio, C., (2013). Ascorbic acid on oral microbial growth and biofilm formation. Pharma Innovation 2, 104-109.
- Zhang, H.M., Wakisaka, N., Maeda, O., Yamamoto, T., (1997). Vitamin C inhibits the growth of a bacterial risk factor for gastric carcinoma: Helicobacter pylori. Cancer 80, 1897-1903.
CrossRef - Vilchèze, C., Hartman, T., Weinrick, B., Jacobs, W.J., (2013). Mycobacterium tuberculosis is extraordinarily sensitive to killing by a Vitamin C‑induced Fenton reaction. Nature Communication 4, 1881.
CrossRef - Bala, I., Bhardwaj, V., Hariharan, S., Kumar, M.N.V.R., (2006). Analytical methods for assay of ellagic acid and its solubility studies. Journal of Pharmaceutical and Biomedical Analysis 40(1), 206-210.
CrossRef

This work is licensed under a Creative Commons Attribution 4.0 International License.








