

Introduction
High fructose and saturated fatty acid-diet reported that increase level of plasma triglyceride and cholesterol in the rats.
High fructose and saturated fatty acid-diet have been reported to increase level of plasma triglyceride and cholesterol in the rats.
High-fructose and/or high-fat diet decrease in TG clearance so induce hypertriglyceridemia.1 In the circulation, the triglycerides carried in chylomicrons are lipolyzed by lipoprotein lipase in the capillary bed of tissues using fatty acids for energy (i.e.skeletal muscle and heart) or storage (i.e.adipose tissue). The fatty acids storage will cause adipose tissue expansion and an increase in fatty acid release into the circulation,2 and activate inflammatory pathways.3,4
Free fatty acids could act as signal molecule activating nuclear factor kappa B (NFκB), an important transcription factor in the inflammatory response.5,4 In this condition, pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-6 and plasminogen activator inhibitor-1 (PAI-1) will increase, whereas the anti-inflammatory cytokines such as IL-10 will decrease.6,7 Reduction of IL-10 expression will increase the severity of inflammation due to the function of this cytokine to inhibit the pro-inflammatory effect.8,9
Interleukin-10 is a pleiotropic cytokine produced and secreted mainly by macrophages (M2).10,11 This cytokine has immuno suppressive roles which inhibits the expression of inflammatory genes.12 The mechanism of action for IL-10 is by inhibiting the activation of NFκB through the kB-inhibitor (IkB) which induces translocation p50 thereby inhibiting the translocation of the heterodimer p65/p50.13 Enhancement of IL-10 expression in an inflammatory state will reduce pro-inflammatory cytokines and reduce the negative effects of inflammation.11
Some studies reported that short chain fatty acid (SCFA) can regulate the production of inflammatory mediators through macrophages.14,15 The SCFA, mainly acetate, propionate, and butyrate, could be produced by colonic bacteria as a result of dietary fiber fermentation. Dietary fiber is an undigested carbohydrate that are resistant to gastrointestinal enzymes but could be fermented by colonic bacteria.16 Propionate acid plays an important role in inhibiting TNF-α and IL-6 secretion and increasing pro-inflammatory cytokines secretion (IL-4 and IL-10).17 Danuyanti et al.,18 reported that high fiber diet decrease TNF-α and IL-6 levels that may be related with suppression of toll-like receptor 4 (TLR4) and NFκB gene expression in hypertriglyceridemia rats. In this study we evaluate the benefits of diet with high fiber gene expression and levels of IL-10 cytokine in white adipose tissue in rats with high fat and fructose diet.
Material and Methods
This study was done after approvement from Ethical Committee of Integrated Research and Testing Laboratory, University of Gadjah Mada (Approval Number: 00065/04/LPPT//2017). Twenty-five (25) male Wistar rats, aged 8 weeks, body weight 180-200 g were divided into 5 groups: 1) normal control rats (N); 2) hyperlipidemia control rats (HL); 3) hyperlipidemia rats with fiber 1.04 g/rat/day (HL1); 4) hyperlipidemia rats with fiber 2.07 g/rat/day (HL2), and 5) hyperlipidemia rats with fiber 3.11 g/rat/day (HL3). The condition of hyperlipidemia was carried out using a diet high in fat and fructose for 7 weeks, and they were considered hyperlypidemia, if their plasma triglyceride levels were > 70.79 mg / dL.19 The blood samples used to analysis triglyceride levels were collected from the medial canthus sinus orbitalis from fasting rats during 10 hours. To get the plasma sample, the blood enters into EDTA tube, and then centrifuge 3000 rpm for 15 minutes. Diet high in fat and fructose were made by replacing fructose into sucrose, and trans-fat into corn starch in the same weight.20,21
The rats were individually in cages and maintained under standard conditions (12:12-h light/dark cycle and 22-25°C room temperatures). Acclimation of the rats was done for 7 days using AIN-93M formulation with modification (L-cystine was substituted by DL-methionine and choline bitartrate by choline chloride) and water ad libitum.22 The normal diet composition consists of 61.94% corn starch, 14% casein, 10% sucrose, 4% corn oil, 5% cellulose, 3.5% mineral mixture, mixture of vitamin 1%, DL-methionine 0.3%, choline chloride 0.25%, and tetrabutilhydroquinone 0.008%. While the treatment diet for T1, T2, and T3 refers to a normal diet with substitution of corn starch using sweet potatoes and pumpkin with a total fiber of 6.88 g, 13.77 g and 20.65 g, respectively per 100 g of diet. The diet fiber content was examined by the Center for Food and Nutrition Studies, University of Gadjah Mada.
Before and after intervention, blood samples were collected from the medial canthus sinus orbitalis for serum IL-10 analysis using ELISA method (FineTest, Wuhan, China) and used as manufacturer’s protocol.23 IL-10 were calculated based on a standard curve and the ELISA assay was performed duplicately. Range detection 31.25-2000pg/mL, the inter-assay: CV<10%, and the intra-assay: CV<8%. White the adipose tissue samples were collected from the retroperitoneal area for IL-10 gene expression analysis using quantitative PCR (qPCR).
The white adipose tissue samples were collected from the retroperitoneal area for IL-10 gene expression analysis by qPCR. Total RNA was extracted using TRIzol reagent (Invitrogen, USA), according to manufacturer’s protocol. Reverse transcription of 1 µg RNA was done based on Revert Aid First Strand cDNA Synthesis Kit (Thermo Scientific, USA). The qPCR assay used SsoFast EvaGreen Supermix (Bio-rad, United Kingdom) with total reaction for qPCR was 10 µL . The conditions of qPCR were 95oC for 10 minutes for early denaturation, followed by 40 cycles of 95oC for 15 sec, 57oC for 1 minute and 72oC for 45 sec.
The conditions of qRT-PCR were 95oC for 10 minutes for early denaturation, followed by 40 cycles of 95oC for 15 sec, 57oC for 1 minute and 72oC for 45 sec. The results were normalized to β-actin housekeeping gene, and relative gene quantification was performed by using the 2− method.24 The specific primer sequences for IL-10 (purchased from Integrated DNA Technologies, Inc, Singapore) were Forward 5’-TTCCCTGGGAGAGAAGCTGA-3’, and Reverse 5’-ATGGCCTTGTAGACACCTTTGT-3’,25 while it for β-actin gene were Forward 5’-ACGGTCAGGTCATCACTATCG-3`, and Reverse 5’-GGCATAGAGGTCTTTACGGATG-3’.26
Statistical Analysis
All data were presented as the mean ± standard deviation (SD). Paired t-tests were used to evaluate the levels of serum IL-10 before and after intervention of high fiber diet. One way ANOVA was used to analyze the differences in IL-10 serum levels and the expression of IL-10 genes between the groups. Tukey’s honest significant difference (HSD) was post hoc tests. Differences were considered statistically significant at p<0.05.
Tukey’s honest significant difference (HSD) was post hoc tests. Differences were considered statistically significant at p<0.05.
Results
The levels of serum IL-10 before and after intervention of high fiber diet to hyperlipidemic rats are shown in Table 1. Before intervention, the levels of IL-10 in the N group were higher than the HL or HL with high-fiber diet groups. It showed that high trans-fat and fructose diet decrease IL-10 levels, which may induce inflammation. However, after administration of the high-fiber diet, the serum levels of IL-10 in hyperlipidemic rats increased significantly (p<0.05). In the present study, there was a decreased serum IL-10 levels in normal control.
Table 1. Mean of IL-10 level before and after high fiber diet.
Table 1: Analysis of serum IL-10 s using ELISA method before and after high fiber diets.
| Group | n | Cytokine level of serum IL-10 (pg/mL) | ∆ Mean (pg/mL) | ∆ Mean (%) | p# | |
| Pre test | Post test | |||||
| Normal (N) | 4 | 100.72±9.74 | 81.87±7.16 | -18.85 | 17.87 | 0.042# |
| Hyperlipidemia (HL) | 4 | 77.81±9.43a | 104.21±10.29a | 26.40 | 33.92 | 0.016# |
| Hyperlipidemia + diet containing 6.88 % fiber (HL1) | 4 | 65.47±9.49a | 123.83±2.75b | 58.36 | 89.14 | 0.001# |
| Hyperlipidemia + diet containing 13.77 % fiber (HL2) | 4 | 74.79±6.97a | 130.65±4.72b | 55.86 | 74.68 | 0.001# |
| Hyperlipidemia + diet containing 20.65% fiber (HL3) | 4 | 76.80±9.95a | 153.50±12.16c | 76.70 | 99.86 | 0.002# |
| p* | 0.251 | 0.001* | ||||
Note: Data is displayed as mean±standard deviation (SD); * p<0.05 is categorized as significant value; # p<0.05 is categorized as significant value; There were no significant differences in pre-test between groups p>0.05. Superscript a,b, and c indicate p<0.05 vs hyperlipidemia group according to One Way ANOVA test followed by Tukey HSD. P in row indicate the differences of serum IL-10 before and after high fiber diet in the same group. P in last row indicates the differences of plasma/serum IL-10 between group. b, c stated p<0.05 vs Hyperlipidemia group.
This study showed that the diet with 20.65% fiber could significantly increase IL-10 gene expression in HL3 (p<0.05). Although the diet containing 13.77% fiber can also increase IL-10 gene expression in HL2, it was not statically significant. It showed that high fiber diet could improve decrease inflammation through increase the gene expression and levels of IL-10. (Figure 1).
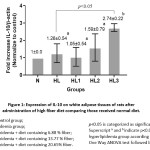 |
Figure 1: Expression of IL-10 on white adipose tissues of rats after administration of high fiber diet comparing those received normal diet. Click here to View figure |
Discussion
In this study, the rats consumption of a high-fat and fructose diet for 7 weeks, in the pre-test, had lower serum IL-10 levels compared to a normal diet (Table 1). This showed that the diets high in fat and fructose could induce a decrease in IL-10 anti-inflammatory cytokines. High fat diet is reported to induce systemic chronic low-grade inflammation.27
High fat diet increases free fatty acid levels that may directly act on intestinal cells, and lead to elevated production of pro-inflammatory27 or decrease anti-inflammatory cytokines. High plasma FFAs could upregulate the expression of TLRs in circulating macrophages, enabling macrophages to be activated (M1 phenotype).27 M1 macrophages secrete TNF-α, IL-1 and MCP-1 to recruit monocytes to adipose tissue and increase the ratio of M1: M2 macrophages that cause a decrease in IL-10 cytokines. The decrease in M2 macrophages can reduce IL-10 levels because M2 macrophages in adipose tissue are important in secreting anti-inflammatory cytokines, especially IL-10, and are involved in maintaining homeostasis in white adipose tissue.28 Therefore, high fat diet causes an increase in proinflammatory cytokines and a decrease in IL-10 levels (Kondo et al., 2018).29, 25, 26
On the other hand, FFA molecules can act as signal transducer molecules that can bind to TLR 2 and TLR 4. Activation of TLR2 and TLR4 will trigger activation of NFkB which causes the release of proinflammatory cytokines and decreased IL-10 levels.7,6 According to Zhu et al.,30 FFA can increase regulation of TLR2 and TLR 4 gene expression when triglycerides (TG), total cholesterol (TC) or both increased.
In this study, both IL-10 gene expression and IL-10 levels in hyperlipidemia rats group were higher than those in normal control rats group. It may be an effect of adipose tissue compensatory. According to Juge-Aubry et al.,31 the adipose tissues always produces anti-inflammatory factors to limit pro-inflammatory effects. IL-10 is one anti-inflammatory factor that will be enhanced to limit TNF-α response during compensation. During acute inflammation, release of pro-inflammatory molecules such as IL-1 and TNF-α is followed release of local anti-inflammatory mediators such as IL-10 as act offset.32 Stoecklin et al.,33 stated that increased expression of IL-10 mRNA was thought to be due to 3-5 h after an increase in TNF-α mRNA. The TNF-α may activate IL-10 gene expression in monocytes.
Administration high fiber diet for 6 weeks was able to increase significantly IL-10 levels in hyperlipidemia rats (HLT1, HLT2, HLT3), p<0.05. Fiber could not be digested by the small intestine enzymes but it would be fermented by colonic bacteria to produce short-chain fatty acids (SCFAs), especially butyrate, propionate and acetate. Butyric and propionate acids were reported to have anti-inflammatory activity.34,35 Sasaki et al.,36 reported that adequate amounts of fiber would increase the fermentation yield of SCFA, for instance, butyric acid. The study by Nastasi et al.,37 showed that butyrate could suppress TNF-α, IL-6 via monocytes and macrophages and may increase the release of IL-10 anti-inflammatory cytokines.
In hyperlipidemia rats, the increase of IL-10 relate to elevate IL-10 gene expression. SCFAs have a possibility to modulate the expression of the IL-10 gene at the mRNA transcription stage. This mechanism accordances with research by Astakhova et al.,38 found that SCFA could limit the activity of HDAC in the nucleus. The inhibition of HDAC activity will give access for transcription factors to the promoter and will provoke gene expression. Besides the effect of high fiber diet intervention, increased IL-10 serum levels in the intervention groups may also be supported by compensatory mechanisms after induction of high trans-fat and fructose. However, the increased IL-10 serum levels in the intervention groups between 2x until 3x compared to the hyperlipidemia rat group without intervention. Therefore, it showed that high-fiber diet could increase IL-10 serum levels.
This study suggested that a high-fiber diet may act as an anti-inflammatory agent, by increasing level and IL-10 gene expression in adipose tissue triggered by high-fat and fructose diets. Based on this study, high-fiber diet could be used to alternative therapeutic strategies in cases of hyperlipidemia and other cases involving inflammatory mechanisms such as obesity, diabetes mellitus and lipid-related cardiovascular disease. In this study, we did not perform the type of dietary fiber. The limitation of this study as we did not perform the analysis of dietary fiber type.
Conclusion
High fiber diet could improve inflammation through increase the gene expression and levels of IL-10 in white adipose tissue of hyperlipidemia rats model.
Acknowledgement
The authors give thanks to Dianandha Septiana Rubi, Budi Lestari and Sukamto for assistance with ELISA, qPCR, and statistical analyses. We are thankful to Klinik Bahasa staff for grammar assistance in writing the manuscript.
Funding
The research was funded by the Directorate General of Higher Education (DIKTI) and Baiturrahmah University Islamic Foundation.
Conflict of Interest
The authors declare there is no conflict of interest to disclose.
References
- Tranchida F., Tchiakpe L., Rakotoniaina Z., Deyris V., Ravion O., Hiol A. Long-term high fructose and saturated fat diet affects plasma fatty acid profile in rats. Journal of Zhejiang University-SCIENCE B. 2012;13 (4): 307-317.
- Ebbert J.O., and Jensen M.D. Fat Depots, Free Fatty Acids,and Dyslipidemia. Nutrients. 2013; 5 (2):498-508.
- Salam D.A., Surya A.S., Tomy D.V., Carla B., Kumar A., Suni S. A review of hyperlipidemia and medicinal plants. International Journal of Applied pharmaceutical Sciences and Biological Sciences. 2013; 2 (4): 219-237.
- Liang H, Lum H, Alvarez A, Garduno-Garcia JJ, Daniel BJ4, Musi N. A low dose lipid infusion is sufficient to induce insulin resistance and a pro-inflammatory response in human subjects. PLoS One. 2018; 13(4): e0195810.
- Brown J. D., Lin C.Y., Duan Q., Griffin G., Federation A., Paranal R.M., Bair S., Newton G., Lichtman A., Kung A., Yang T., Wang, H., Luscinskas, F.W., Croce K., Bradner, J.E., Plutzky, J. NF-kB directs dynamic super enhancer formation in inflammation and atherogenesis. Molecular Cell. 2014; 56 (2): 219–231.
- Fritsche K.L. The science of fatty acids and inflammation. Advances in Nutrition. 2015; 6(3): 293–301.
- Charles B.A., Doumatey A., Huang H., Zhou J., Chen G., Shriner D., Adeyemo A., Rotimi C.N. The roles of IL-6, IL-10, and IL-1ra in obesity and insulin resistance in African-Americans. J Clin Endocrinol Metab. 2011; 96 (12): 2018–2022.
- Driessler F., Venstrom K., Sabat R., Asadullah K., Schottelius A. J. Molecular mechanisms of interleukin-10-mediated inhibition of NF-kB activity: a role for p50. Clin Exp Immunol. 2004; 135 (1): 64–73.
- Bazzoni F., Tamassia N., Rossato M., Cassatella M.A. Understanding the molecular mechanisms of the multifaceted IL-10-mediated anti-inflammatory response: lessons from neutrophils. Eur J Immunol. 2010; 40 (9): 2360-2368.
- Shouval D.S., Biswas A., Goettel J.A., McCann K., Conaway E., Redhu N.S., Mascanfroni I.D., Al Adham Z., Lavoie, S., Ibourk M., Nguyen D.D., Samsom J.N., Escher J.C., Somech R., Weiss B., Beier R., Conklin L.S., Ebens C.L., Santos F.G., Ferreira A.R., Sherlock M., Bhan A.K., Müller W., Mora J.R., Quintana F.J., Klein C., Muise A.M., Horwitz B.H., Snapper S.B. Interleukin-10 receptor signaling in innate immune cells regulates mucosal immune tolerance and anti-inflammatory macrophage function. Immunity. 2014; 40 (55): 706–719.
- Xie L., Fu Q., Ortega T.M., Zhou L., Rasmussen D., O’Keefe J., Chapes S.K. Overexpression of IL-10 in C2D macrophages promotes a macrophage phenotypic switch in adipose tissue environments. PLoS ONE. 2014; 9 (1):
- Antoniv T.T., Ivashkiv L.B. Interleukin-10-induced gene expression and suppressive function are selectively modulated by the PI3K-Akt-GSK3 pathway. Immunology. 2011; 132 (4): 567-77.
- Andersen V., Egeberg R., Tjønneland A., Vogel U. Interaction between interleukin-10 (IL-10) polymorphisms and dietary fibre in relation to risk of colorectal cancer in a Danish case-cohort study. BMC Cancer. 2012; 12 (183): 1-9.
- Weickert M.O., Pfeiffer A.F.H. Metabolic effects of dietary fiber consumption and prevention of diabetes. The Journal of Nutrition. 2008; 138 (3): 439–442.
- Kim K.A., Gu W., Lee I.A., Joh E.H., Kim D.H. High fat diet-induced gut microbiota exacerbates inflammation and obesity in mice via the TLR-4 signaling pathway. 2012; PLoS ONE; 7 (10): e47713.
- Cummings J.H., Pomare E.W., Branch W.J., Naylor C.P.E., Macfarlane G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut; 1987; 28 (10):1221- 1227.
- Al-Lahham S.H., Peppelenbosch, M.P., Roelofsen H., Vonk R.J., Venema K. Biological effects of propionic acid in humans; metabolism, potential applications and underlying mechanisms. Biochim Biophys Acta. 2010; 1801 (11): 1175-1183.
- Danuyanti I.G.A.N., Farmawati A., Sunarti. High Fiber Diet Suppression of TLR4 and NFκβ Gene Expression Correlated with TNF-α and IL-6 Levels in Hypertriglyceridemia Rats. Journal of Food and Nutrition Research. 2018; 6 (12): 725-730.
- Ihedioha J.I., Noel-Uneke O.A., Ihedioha T.E. Reference values for the serum lipid profile of albino rats (Rattus norvegicus) of varied ages and sexes. Comp Clin Pathol. 2013; 22 (1): 93-99.
- Ble-Castillo J.L., Aparicio-Trapala M.A., Juárez-Rojop I.E., Torres-Lopez J.E., Mendez J.D., Aguilar-Mariscal H., Olvera-Hernández V., Palma-Cordova L.C., Diaz-Zagoya J.C. Differential effects of high-carbohydrate and high-fat diet composition on metabolic control and insulin resistance in normal rats. Int J Environ Res Public Health. 2012; 9 (5): 1663-1676.
- Sasidharan S.R., Joseph J.A., Anandakumar S.,Venkatesan V., Madhavan C.N.A., Agarwal A. An experimental approach for selecting appropriate rodent diets for research studies on metabolic disorders. BioMed research international. 2013; 2013: 1-9.
- El-sheikh, N., El Fattah H.M.A. Counteracting Methionine Choline-Deficient Dietinduced Fatty Liver by Administration of Turmeric and Silymarin. J Appl Sci Res. 2011; 7 (12): 1812-1820.
- Putra A, Ridwan FB, Putridewi AI, Kustiyah AR, Wirastuti K, Sadyah NAC, Rosdiana I, Munir D. The Role of TNF-α induced MSCs on Suppressive Inflammation by Increasing TGF-β and IL-10. Open Access Maced J Med Sci. 2018; 6(10):1779-1783.
- Livak K.J., Schmittgen T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCt Method. METHOD. 2001; 26: 402-408.
- Gonzalez P, Burgaya F, Acarin L, Peluffo H, Castellano B, Gonzalez B. Interleukin-10 and interleukin-10 receptor-I are upregulated in glial cells after an excitotoxic injury to the postnatal rat brain. J Neuropathol Exp Neurol. 2009; 68(4):391-403.
- Feng K, Ge Y1, Chen Z1, Li X1, Liu Z1, Li X1, Li H2, Tang T1, Yang F1, Wang X1. Curcumin Inhibits the PERK-eIF2α-CHOP Pathway through Promoting SIRT1 Expression in Oxidative Stress-induced Rat Chondrocytes and Ameliorates Osteoarthritis Progression in a Rat Model. Oxid Med Cell Longev. 2019; 2019:8574386.
- Duan Y., Zeng L., Zheng C., Song B., Li F., Kong X., and Xu K. Inflammatory Links Between High Fat Diets and Diseases. Front Immunol. 2018; 9: 2649.
- Castoldi A., Naffah de Souza C., Câmara N.O., Moraes-Vieira P.M. The macrophage switch in obesity development. Frontiers in Immunology. 2016; 6 (637):1-7.
- Kondo H, Abe I, Gotoh K, Fukui A, Takanari H, Ishii Y, Ikebe Y, Kira S, Oniki T, Saito S, Aoki K, Tanino T, Mitarai K, Kawano K, Miyoshi M, Fujinami M, Yoshimura S, Ayabe R, Okada N, Nagano Y, Akioka H, Shinohara T, Akiyoshi K, Masaki T, Teshima Y, Yufu K, Nakagawa M, Takahashi N. Interleukin 10 Treatment Ameliorates High-Fat Diet-Induced Inflammatory Atrial Remodeling and Fibrillation. Circ Arrhythm Electrophysiol. 2018; 11(5):e006040.
- Zhu Y.J., Wang C., Song, G., Zang S.S., Liu Y., Li L. Toll-like receptor-2 and -4 are associated with hyperlipidemia. Molecular Medicine Reports. 2015; 12 (6): 8241-8246.
- Juge-Aubry C.E., Somm E., Pernin A., Alizadeh N., Giusti V., Dayer J.M., Meier C.A. Adipose tissue is a regulated source of interleukin-10. Cytokine. 2005; 29 (6):270-274.
- Stumpo,D., Lai W.S., Blackshear P.J. Inflammation: cytokines and RNA-based regulation. Wiley Interdiscip Rev RNA. 2010; 1 (1): 60–80.
- Stoecklin G., Tenenbaum S.A., Mayo T., Chittur S.V., George AD., Baroni T.E., Anderson P. Genome-wide analysis identifies interleukin-10 mrna as target of tristetraprolin. J Biol Chem. 2008; 283 (17): 1689–1169.
- Canani R.B., Di-Costanzo M., Leone L., Pedata M., Meli R., Calignano A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J Gastroenterol. 2011; 17 (12): 1519-1528.
- Besten G.D., Eunen K.V., Groen A.K., Venema K., Reijngoud D.J., Bakker B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013; 54 (9):2325-40.
- Sasaki D., Sasaki K., Ikuta N., Yasuda T., Fukuda I., Kondo A., Osawa R. Low amounts of dietary fibre increase in vitro production of short-chain fatty acids without changing human colonic microbiota structure. Scientific Report. 2018; 8 (435): 1-9.
- Nastasi C., Candela M., Bonefeld C.M., Geisler C., Hansen M., Krejsgaard T., Biagi E., Andersen M.H., Brigidi P., Ødum N., Litman T., Woetmann A. The effect of short-chain fatty acids on human monocyte-derived dendritic cells. Scientific Reports. 2015; 5 (16148): 1-10.
- Astakhova L., Ngara Mi., Babich O., Prosekov A., Asyakina L., Dyshlyuk L, Midtvedt T., Zhou X., Ernberg I., Matskova L. Short chain fatty acids (SCFA) reprogram gene expression in human malignant epithelial and lymphoid cells. PLoS ONE. 2016; 11 (7): e0154102.

This work is licensed under a Creative Commons Attribution 4.0 International License.








